Pantoteenihappo
-Pantothenic_acid_Formula_V.1.svg)
| |

| |
Pantoteenihappo
| |
| Systemaattinen (IUPAC) nimi | |
| 2-[[(2R)-2,4-dihydroksi-3,3-dimetyylibutanoyyli]amino]propaanihappo | |
| Tunnisteet | |
| CAS-numero | |
| ATC-koodi | A11 |
| PubChem CID | |
| DrugBank | |
| Kemialliset tiedot | |
| Kaava | C9H17NO5 |
| Moolimassa | 219,237 g/mol |
| Fysikaaliset tiedot | |
| Tiheys | Ca2+-suola:[1] 1,32 g/cm³ |
| Sulamispiste | 195–196 °C (Ca2+-suola)[3] |
| Liukoisuus veteen | Hyvin liukoinen (pantoteenihappo ja sen monet suolat)[2] |
| Farmakokineettiset tiedot | |
| Hyötyosuus | 40–60 % (ruuasta)[4] |
| Metabolia | ~15 % hapettuu energiaksi[4] |
| Puoliintumisaika | ? |
| Ekskreetio | Virtsa[4] |
| Terapeuttiset näkökohdat | |
| Raskauskategoria |
A(US); C(US) jos >6–7 mg/vrk[5] |
| Reseptiluokitus | |
| Antotapa | nieltynä[5] |
Pantoteenihappo eli B5-vitamiini on pantoiinihappo, johon on liittynyt β-alaniini amidisidoksella. Pantoteenihappo on välttämätön vitamiini ihmisille ja lähes kaikille muille eliöille. Melkein kaikki kasvit, bakteerit ja arkit valmistavat sitä, mutta mitkään eläimet eivät. Sitä saadaan kylliksi lähes kaikista ruuista ja sen puutos on ihmisillä erittäin harvinainen tila.[6]
Eliöt muuntavat pantoteenihappoa koentsyymi A:ksi (CoA), joka kiinnittyy tiettyihin eliöiden entsyymeihin entsyymitoiminnan kannalta pakollisena molekyylinä eli koentsyyminä.[6] Eliöt muuntavat CoA:ta myös kantajaproteiineihin, kuten asyylinkantajaproteiineihin, kovalenttisesti sitoutuneiksi prosteettisiksi ryhmiksi.[4][7]
Pantoteenihappo on CoA:n ja prosteettisten ryhmien muodossa osa satoja eri entsyymityyppejä. Joidenkin näiden entsyymien avulla ihmiset ja monet muut eliöt hajottavat hiilihydraatteja, aminohappoja ja rasvahappoja energiaksi eli ATP:ksi (katso sitruunahappokierto, ureakierto ja β-oksidaatio). Pantoteenihappo on myös osa rasvahappojen ja steroidien valmistusta sekä asetylaatioreaktioiden kautta osana muun muassa asetyylikoliinin ja asetyloitujen proteiinien tuottoa.[4]
Pantoteenihapon anioni on pantotenaatti.
Suositukset ja saanti[muokkaa | muokkaa wikitekstiä]
Suomen valtion ravitsemusneuvottelukunta ei ole esittänyt ravitsemussuosituksia pantoteenihapolle.[8]
EFSA ei myöskään ole esittänyt saantisuosituksia pantoteenihapolle tutkimustiedon puutteen vuoksi. EFSA on kuitenkin esittänyt arvot riittävälle saannille eli AI-arvot (eng. adequate intake). Nämä ovat arvioita siitä, mikä määrä pantoteenihappoa riittää ylläpitämään jonkin väestöryhmän terveenä. Ryhmät on eritelty alla olevassa taulukossa.[9] Eräs ero todellisen vähimmäistarpeen keskiarvon ja AI-arvon välillä on se, että AI-arvon tulisi olla suurempi.[10]
| Ikä | Miehet
ja naiset |
Raskaana
olevat |
|---|---|---|
| 7–11 kk | 3 | 5 |
| 1–3 v | 4 | |
| 4–6 v | 4 | |
| 7–10 v | 4 | Imettävät |
| 11–14 v | 5 | 7 |
| 15–17 v | 5 | |
| ≥18 v | 5 |
Liikasaanti[muokkaa | muokkaa wikitekstiä]
EFSA ei ole asettanut pantoteenihapon päiväsaannille ylärajaa.[12] Se on verrattain turvallinen aine ja keho rajoittaa sen imeytymistä.[6] Ihmisillä 10–15 gramman vuorokausiannokset pantoteenihapon kalsiumsuolaa eivät ole noin vuoden jatkuneen kuurin aikana aiheuttaneet kuin väliaikaista ja lievää pahoinvointia sekä vatsakipuja.[13] Myöskään joissain B5-vitamiinilisissä käytetty pantenoli ei ole myrkyllinen.[6]
Pantoteenihapon LD50-arvot suun kautta hiirille ja rotille ovat 10 grammaa per kehonpainokilogramma (g/kg), jolloin kuoleman aiheuttaa hengityskatko. Rotille 6 kuukautta jatkunut 500 tai 2 000 milligramman pantoteenihappoannosten syöttö per kehonpainokilogramma joka vuorokausi (mg/kg/vrk), 50 mg/kg/vrk koirille ja 200–250 mg/kg/vrk apinoille eivät ole aiheuttaneet kyseisillä eläimillä näkyviä tai ruumiinavauksessa havaittavia haittoja.[6]
Puutos[muokkaa | muokkaa wikitekstiä]
Pantoteenihapon puutos ihmisissä ja muissa eliöissä on hyvin harvinainen tila, sillä pantoteenihappoa on monissa ruuissa kylliksi suhteessa päivittäiseen tarpeeseen. Puutoksen oireet vaihtelevat eliöstä toiseen.[4]
Ihmisillä[muokkaa | muokkaa wikitekstiä]
Ihmisillä pantoteenihappopuutoksen oireita on havaittu lähinnä vain koetilanteissa. Pantoteenihappovajaa ruokavalio on noin 9 viikon kuluttua aiheuttanut väsymystä terveillä koehenkilöillä.[14] Oireita on saatu aikaan myös ihmisillä, jotka ovat tutkimuksissa saaneet pantoteenihapon toimintaa estäviä aineita eli antagonisteja (katso kohta antagonistit). Näillä saavutetun puutoksen oireita ovat olleet esimerkiksi poltteleva kipu jalkaterissä, masennus, fyysinen uupumus, unettomuus, oksentelu, lihasheikkous, ruuansulatusongelmat, insuliiniresistenssin väheneminen ja vasta-aineiden tuoton väheneminen.[4]
Nälkiintyneillä ihmisillä, kuten 2. maailmansodan sotavangeilla, on joissain tapauksissa ilmennyt muun muassa jalkojen polttelua ja tunnottomuutta, jota kutsutaan nykyään Grierson-Gopalan oireyhtymäksi. Näiden vankien oireiden esitetään usein johtuneen pantoteenihapon puutteesta eritoten Colothur Gopalan tutkimusten vuoksi.[15][16] Hän havaitsi polttelun lakkaavan vain pantoteenihappokuurilla,[15] mutta myöhemmät kokeet eivät ole tukeneet Gopalan johtopäätöstä siitä, että oireet johtuvat pantoteenihapon puutoksesta.[17][16]
Puutos voi myös olla geneettinen. Pantotenaattikinaasi-entsyymin (EC-numero 2.7.1.33) muodon 2 mutaatiosta johtuva toimimattomuus estää kehoa tuottamasta kylliksi koentsyymi A:ta pantoteenihaposta. Kinaasin muut muodot (muodot 1, 3 ja 4) kompensoivat tätä puutosta, mutta se johtaa silti vuonna 1924 löydettyyn hyvin harvinaiseen ja kuolettavaan tautiin,[18] jota sanotaan nimellä pigmentoitunut pallidusrappeuma, joka on englanniksi pantothenate kinase-associated neurodegeneration eli PKAN.[19][18] Tämän oireita ovat muun muassa näköhäiriöt ja raudan kertyminen aivoihin.[18]
Muilla eläimillä[muokkaa | muokkaa wikitekstiä]
Rotilla pantoteenihappopuutos aiheuttaa muun muassa ihottumaa, turkin lähtöä, turkin harmaantumista, lisämunuaisten kuolion, pohjukaissuolen haavautumista, kouristelevan liikkumistavan, anemiaa, leukopeniaa, vasta-aineiden tuoton vähentymistä, sukupuolirauhasten surkastumista ja hedelmättömyyttä. Koirilla puutos voi johtaa ruokahaluttomuuteen, ripuliin, akuuttiin aivovaurioon, koomaan, hypoglykemiaan, rasvamaksaan, mitokondrioiden koon kasvuun, veren valkosolumäärän kasvuun (leukosytoosi), veren ammoniakkiapitoisuuden kasvuun (hyperammonemia) ja veren laktaattipitoisuuden kasvuun (hyperlaktemia). Sioilla oireita ovat muun muassa ihottuma, ripuli, kyynelehtiminen, haavainen paksusuolen tulehdus ja selkäydinhermon ja ääreishermoston vaurioituminen, joihin liittyy kouristeleva kävelytapa. Kanoilla puutos voi aiheuttaa ihottumaa nokan, silmien ja jalkojen alueella. Voi myös ilmetä muun muassa sulkien kasvun hidastumista, selkäydinhermon myeliinin tuhoutumista, kateenkorvan surkastumista ja rasvamaksan kehittyminen.[6]
Ruokapitoisuudet[muokkaa | muokkaa wikitekstiä]
Pantoteenihappoa lähes kaikissa ruuissa. Sitä on paljon eläinten lihoissa ja eritoten sisäelimissä, kuten maksoissa ja sydämissä. Sitä on paljon myös joissain viljoissa, pähkinöissä, sienissä ja kasviksissa. Luonnossa kaikkein eniten pantoteenihappoa on joidenkin kylmän veden kalojen kuten tonnikalan ja turskien munasarjoissa (yli 2,3 milligrammaa per gramma ruokaa, mg/g) ja hunajamehiläisten kuningatarhyytelössä (0,5 mg/g).[4]
Täysjyväviljoissa pantoteenihappo on pääosin kuoriosissa, kuten leseissä, joten niiden poistaminen vähentää viljan pantoteenihappopitoisuutta. Pitkäaikainen säilytys huoneenlämmössä ja ruuanlaitossa tavallisesti käytetyt lämpötilat eivät tuhoa sitä merkittävästi. Lihan paistossa pantoteenihappohäviö kuitenkin on 15–50 % ja kasvisten kuumentamisessa häviö on 37–78 %. pH-arvossa, joka on yli 7 tai alle 5, pantoteenihappo tuhoutuu helposti. Maidon pastöroinnissa häviö ei ole suuri, sillä maito on lähes pH-neutraali.[4] Säilyketölkkipakkaaminen voi johtaa ruuasta ja pakkausmenetelmistä riippuen esimerkiksi 30–80 % häviöön.[20] Pakastaminen ei tuhoa pantoteenihappoa, mutta sulatusvesien mukana suuri osa siitä voi poistua sen vesiliukoisuuden takia.[21]
Ihmisillä ruokien pantoteenihaposta noin 40–60 % imeytyy suolistosta. Aine on ruuissa pääosin sitoutunut koentsyymi A:han tai proteiineihin.[4]
| Heinäkasvit ja jauhot | Pavut ja pähkinät | Lihat | |||
|---|---|---|---|---|---|
| Kaurahiutale | 1,349 | Cashewpähkinä, kuivapaahdettu | 1,217 | Ankka, paahdettu | 1,5 |
| Maissijauho, täysjyvä | 0,658 | Härkäpapu, keitetty | 0,157 | Kalkkuna, iholla, paahdettu | 0,858 |
| Ohraryyni | 0,135 | Kidneypapu, keitetty | 0,22 | Kana, iholla, paahdettu | 0,926 |
| Pasta, täysjyvä, keitetty | 0,419 | Kikherne, keitetty | 0,286 | Kirjolohi (viljelemätön), paistettu | 1,065 |
| Riisi, tumma, keitetty | 0,285 | Linssi, keitetty | 0,638 | Makrilli, paistettu, kuivattu | 0,99 |
| Riisi, valkoinen, keitetty | 0,215 | Maapähkinä, kuivapaahdettu | 1,395 | Nauta (vasikka), maksa, käristetty[23] | 8,8 |
| Ruisjauho, puolikarkea | 0,492 | Manteli, kuivapaahdettu | 0,254 | Nauta, kylki, ¼ rasvaa, pariloitu | 0,32 |
| Soijajauho, rasvainen | 1,209 | Parapähkinä, kuivapaahdettu | 0,236 | Nauta, munuainen, pariloitu[23] | 3,00 |
| Vehnäjauho, täysjyvä | 1,008 | Pekaanipähkinä, kuivapaahdettu | 1,774 | Sika, jalka, vähärasvainen, paahdettu | 0,67 |
| Vehnälese | 2,181 | Pistaasi, kuivapaahdettu | 0,255 | Silli, säilyke | 0,081 |
| Vehnänalkio | 2,257 | Voipapu, keitetty | 0,257 | Turska, paistettu | 0,18 |
| Kasvikset ja ruokasienet | Hedelmät ja marjat | Lehmänmaitotuotteet | |||
| Bataatti, kuorimaton, uunipaistettu | 0,646 | Aitoviikuna | 0,3 | Briejuusto | 0,69 |
| Herne | 0,104 | Ananas | 0,16 | Camembertjuusto | 1,364 |
| Jääsalaatti | 0,046 | Appelsiini | 0,25 | Cheddarjuusto | 0,413 |
| Keltasipuli | 0,106 | Aprikoosi | 0,24 | Edamjuusto | 0,281 |
| Keräkaali, keitetty | 0,139 | Avokado | 0,971 | Maito, rasvaton | 0,329 |
| Kukkakaali, keitetty | 0,508 | Banaani | 0,26 | Raejuusto, 1 % rasvaa | 0,215 |
| Kurkku | 0,178 | Hunajameloni | 0,128 | Sinihomejuusto | 1,729 |
| Lanttu | 0,155 | Karviainen | 0,286 | Voi | 0,11 |
| Maissi, keltainen | 0,76 | Luumu | 0,182 | Kerma, 31,3 % rasvaa[23] | 0,26 |
| Parsa, keitetty | 0,161 | Mango | 0,16 | Herajauhe, makea[23] | 5,62 |
| Parsakaali, keitetty | 0,508 | Mansikka | 0,34 | Jugurtti, 3,3 % rasvaa[23] | 0,59 |
| Peruna, kuorimaton, uunipaistettu | 0,555 | Mustaherukka | 0,398 | Muut | |
| Pinaatti | 0,065 | Mustikka | 0,093 | Kananmuna, keitetty | 1,398 |
| Porkkana | 0,197 | Omena | 0,061 | Rintamaito (ihmisen) | 0,011 |
| Punajuuri, keitetty | 0,145 | Persikka | 0,17 | Rypsiöljy | 0 |
| Selleri | 0,186 | Päärynä | 0,07 | Oliiviöljy | 0 |
| Siitake, kuivattu | 21,879 | Vadelma | 0,24 | Olut, lager | 0,058 |
| Tomaatti | 0,247 | Viinirypäle | 0,283 | Leivinhiiva, tuore (puristehiiva)[23] | 3,50 |
| a: ruuat ovat raakoja eli valmistamattomia ellei toisin mainita. Pitoisuudet ovat keskimääräisiä. | |||||
Kemia[muokkaa | muokkaa wikitekstiä]
Pantoteenihapon OH-ryhmän kohdalla on kiraliakeskus, joten se on optisesti aktiivinen ja sillä kaksi enantiomeeriä. Näistä vain D- eli (+)-(R)-pantoteenihappo toimii vitamiinina.[24] (R)-pantoteenihapon CAS-numero on 79-83-4 ja rasemaatin 599-54-2.
(R)-Pantoteenihappo on viskoosi, kellertävä[3] tai väritön hygroskooppinen öljy. Se on hyvin liukoinen veteen ja etanoliin sekä vähäliukoinen dietyylieetteriin. Se on epävakaa happamissa ja emäksisissä liuoksissa,[2] mutta vakaa neutraalissa liuoksessa. Lämmittäminen,[6] UV-valo ja happi edistävät sen hajoamista. Se säilyy parhaiten viileässä ja pH-arvossa 7.[25]
Pantoteenihapolla on hyvin heikko absorbanssimaksimi aallonpituudella 204 nm karbonyylinsä vuoksi.[26] Aine ei fluoresoi.[27]
Suolat[muokkaa | muokkaa wikitekstiä]
Pantoteenihapon suolat ovat vakaampia kuin pelkkä pantoteenihappo. Siksi suoloja käytetään elintarvikelisänä ja lisäravintoaineissa hapon sijaan. Useimmin käytetty suola on kalsiumpantotenaatti.[6]
Kalsium-(R)-pantotenaatin (CAS 137-08-6) massa on 476,5 g/mol.[2] Se on valkea ja hajuton jauhe,[6] lievästi hygroskooppinen, kohtalaisen vakaa ilmassa ja valossa. Se on makea, mutta sillä on hieman kitkerä jälkimaku. Se hajoaa sulamatta 195–196 °C lämpötilassa.[28] Se on hyvin vesiliukoinen (1 g liukenee 2,8 ml vettä 25 °C:ssa),[1] liukoinen glyseroliin, hiukan liukoinen etanoliin ja asetoniin. Sen laimeat vesiliuokset ovat heikosti emäksisiä.[28] Sen tiheys on 1,32 g/ml ja irtotiheys jauheena 0,57–0,67 g/ml.[1]
Natrium-(R)-pantotenaatin (CAS 867-81-2) massa on 241,2 g/mol. Se on hygroskooppinen valkea jauhe, joka hajoaa sulamatta 160–165 °C lämpötilassa. Se on hyvin liukoinen veteen, hieman liukoinen etanoliin (paremmin kuin kalsiumsuola) ja liukenematon dietyylieetteriin.[2]
Johdannaiset[muokkaa | muokkaa wikitekstiä]
Alla on pantoteenihapon kanssa rakenteeltaan ja nimeltään samankaltaisia aineita. Näistä esimerkiksi (R)-pantenolia ei ole luonnossa, mutta se hapettuu silti ihmisissä ja muissa eläimissä aktiiviseksi B5-vitamiiniksi.[2] Pantenoli on vakaampi kuin pantoteenihappo, jonka vuoksi sitä käytetään joskus B5-vitamiinilisissä.[4]
-
 (R)-pantoiinihappo
(R)-pantoiinihappo
CAS 1112-33-0 -
 (R)-pantetiini
(R)-pantetiini
CAS 16816-67-4 -
 (R)-panteteiini
(R)-panteteiini
CAS 496-65-1 -
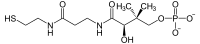 (R)-4'-fosfopanteteiini
(R)-4'-fosfopanteteiini -
 (R)-pantenoli
(R)-pantenoli



Valmistus[muokkaa | muokkaa wikitekstiä]
Pantoteenihappoa valmistetaan teollisesti kemiallisin synteesein. Mikrobimenetelmät ovat kuitenkin kehittyneet lähes kaupallisesti toteuttamiskelpoiselle tasolle.[29] (R)-Pantoteenihapon kalsiumsuolaa valmistetaan reagoimalla esimerkiksi (R)-pantolaktonia ja β-alaniinin kalsiumsuolaa kiehuvassa metanolissa tai etanolissa. Tuote puhdistetaan uudelleenkiteyttämällä ja haihduttamalla.[2]
Toiminta eliöissä[muokkaa | muokkaa wikitekstiä]
Eliöt käyttävät (R)-pantoteenihappoa koentsyymi A:n (CoA:n) valmistukseen. CoA:lla on sellaisenaan käyttötarkoituksia eliöissä, mutta sitä myös kulutetaan erilaisten kantajaproteiinien viimeistelyyn translaation jälkeen 4'-fosfopanteteinyylitransferaasi-entsyymeillä. Näissä CoA:lta siirretään (R)-pantoteenihappo kantajaproteiineihin (R)-4'-fosfopanteteiinina. Tämä liittyy kantajaproteiineihin kovalenttisesti ollen siten niiden prosteettinen ryhmä.[7]
Kantajaproteiinit[muokkaa | muokkaa wikitekstiä]
Kantajaproteiineissa (R)-4'-fosfopanteteiinin tioli (–SH) on vapaa.[7] Se muodostaa väliaikaisia tioesterisidoksia pienten molekyylien kanssa, jotka se siirtää molekyyliltä toiselle (R)-pantoteenihapon taipuisan rungon ansiosta.[6][7] Kantajaproteiineja ovat
- asyylikantajaproteiinit (ACP) – nämä siirtävät asyyleitä ja ovat osa rasvahapposyntaaseita (FAS, EC-numero 2.3.1.85) ja polyketidisyntaaseita (PKS).[7] FAS:iä on mikrobeissa, kasveissa ja eläimissä. FAS:t osallistuvat rasvahappojen valmistukseen.[6] PKS:iä on monissa bakteereissa, sienissä ja kasveissa. PKS:t ovat suuri joukko erikoistuneita entsyymeitä, jotka valmistavat polyketidejä, kuten tetrasykliiniä ja sirolimuusia.[30]
- peptidyylikantajaproteiinit (PCP) – nämä siirtävät peptidejä ja aminohappoja ei-ribosomaalisilla peptidisyntaaseilla (NRPS) tehden ei-ribosomaalisia peptidejä, joita ovat esimerkiksi penisilliinit, bleomysiini, siklosporiini, mikrokystiinit ja nodulariinit.[31] NRPS:iä on bakteereissa ja sienissä, muttei eläimissä ja kasveissa.[32]
- aryylinkantajaproteiinit (ArCP) – nämä siirtävät aryylien karboksyylihappoja, kuten salisylaatteja ja 2,3-dihydroksibentsoaatteja, ja osallistuvat esimerkiksi enterobaktiinin valmistukseen bakteereissa, johon osallistuvat myös NRPS:sät.[33]
Koentsyymi A[muokkaa | muokkaa wikitekstiä]
- Pääartikkeli: Koentsyymi A
CoA on nimensä mukaisesti koentsyymi eli entsyymeitä avustava molekyyli. Entsyymeissä se siirtää asyyliryhmiä molekyyliltä toiselle asyyli-CoA:na. CoA:n adeniiniosa sitoutuu proteiineihin. (R)-4'-panteteiini taas toimii taipuisana "kätenä", jonka rikkiin asyylit sitoutuvat väliaikaisesti tioestereiksi (samalla tapaa kuin kantajaproteiineissa).[6] CoA on satojen erityyppisten entsyymien koentsyymi.[4] Yksi tärkeä esimerkki on sitruunahappokierto, jossa CoA:n siirtämä asyyli on pyruvaatista saatu etikkahappo, eli muodostunut aine on asetyyli-CoA. Kierrossa asetyylin hiiliatomit hapetetaan ilman hapella hiilidioksidiksi. Samalla saadaan soluille ATP:tä.[34]
Muu biokemia[muokkaa | muokkaa wikitekstiä]
Imeytyminen[muokkaa | muokkaa wikitekstiä]
Ruokien pantoteenihaposta pääosa on sitoutunut koentsyymi A:han tai asyylikantajaproteiiniin. Näistä se irtoaa ohutsuolessa 4'-fosfopanteteiiniksi, jota voi olla myös ruuassa jo valmiiksi.[4] Edeltävien aineiden hydrolyysi tapahtuu suoliston pyrofosfataasien ja fosfataasien toimesta, jolloin muodostuu 4'-fosfopanteteiinin kautta pantetiinia. Tämä hajoaa edelleen pantoteenihapoksi pantotenaasilla (EC-numero 3.5.1.22). Suurissa pitoisuuksissa pantoteenihapon imeytyminen suolistosta tapahtuu passiivisesti, mutta sen tyypillisissä ruokapitoisuuksissa imeytyminen tapahtuu energiaa kuluttavasti kalvoproteiinivälitteisesti.[6]
Ihmisillä pantoteenihapon imeytyminen tapahtuu SMVT-kalvoproteiinin kautta (eng. sodium-dependent multivitamin transporter). Tämän geeni on SLC5A6. Myös biotiini imeytyy SMVT:n avulla. SMVT:t ovat symporttereita ja ottavat pantoteenihappoa (ja biotiinia) sisälle soluihin natriumionien siirtyessä samanaikaisesti sisälle soluun.[35]
Kuljetus[muokkaa | muokkaa wikitekstiä]
Ihmisillä pantoteenihappo kulkeutuu veriplasmassa vapaana pantoteenihappona, jota punasolut ottavat sisäänsä passiivisella diffuusiolla. Tästä osan ne muuntavat 4'-fosfopantoteeniksi (CAS 5875-50-3) ja pantetiiniksi. Veren pantoteenihaposta pääosa on punasoluissa. Aikuisen veressä pantoteenihappoa on usein noin 1,12–1,96 µg/ml, josta plasmassa on 0,211–1,96 µg/ml. Vanhuksilla pitoisuus voi olla matalampi, noin 0,5–0,7 µg/ml.[4]
Varastoituminen[muokkaa | muokkaa wikitekstiä]
Ihmiskeho ei varastoi ylimäärin pantoteenihappoa. Sen pitoisuudet eivät silti juuri muutu kehossa rajoitettaessa sen saantia ravinnosta, sillä keho kierrättää sitä tehokkaasti. Ihmisillä pantoteenihappo on kertyneenä kehoon pääosin koentsyymi A:n muodossa. Aikuisella on maksassa noin 28 mg pantoteenihappoa sen eri muodoissa. Ainetta on paljon myös lisämunuaisissa, munuaisissa, aivoissa, sydämessä ja kiveksissä. Sydän- ja maksasoluissa aineesta vastaavasti 95 % ja 70 % on mitokondrioissa.[4]
Hajotus ja erittyminen[muokkaa | muokkaa wikitekstiä]
Ihmisillä pantoteenihappo erittyy pääosin virtsassa pantoteenihappona (noin 0,8–8,4 mg/vrk) ja 4'-fosfopanteteiinina. Normaalin kokoisin annoksin noin 15 % päivittäin saadusta pantoteenihaposta hapettuu energiaksi, eli osa siitä poistuu kehosta hengitysilman hiilidioksidina.[4]
Biosynteesi[muokkaa | muokkaa wikitekstiä]
Lähes kaikki kasvit, bakteerit ja arkit pystyvät tekemään pantoteenihappoa. Ne tekevät sitä pääosin liittämällä pantoiinihappoja ja β-alaniineihin amidisidoksin.[6] Reaktioreitti on alla.

α-Ketovaleraattia eli 3-metyyli-2-oksobutanoaatin anioneja (CAS-numero 1821-02-9) muodostuu (R)-2,3-dihydroksi-isovaleraatista dihydroksihappodehydrataasilla (EC-numero 4.2.1.9) tai L-valiinista BCAT-entsyymillä (EC 2.6.1.42). Kumpi näistä reiteistä on pääasiallinen, riippuu eliöstä.[36]
α-Ketovaleraattiin liittyy hydroksimetyyli (-CH2OH) 3-metyyli-2-oksobutanoaattihydroksimetyylitransferaasilla (EC 2.1.2.11):[36][6]
- α-ketovaleraatti + CH2-tetrahydrofolaatti → 2-dehydropantoaatti + tetrahydrofolaatti
2-Dehydropantoaatti eli pantoiinihapon anioni (CAS 470-30-4) pelkistyy 2-dehydropantoaatti-2-reduktaasilla (EC 1.1.1.169):[36][6]
- 2-dehydropantoaatti + NADPH → (R)-pantoaatti + NADP+
(R)-pantoaatti eli pantoiinihappoanioni (CAS 1112-33-0) liittyy β-alaniiniin pantoaatti-beeta-alaniiniligaasilla (EC 6.3.2.1), jolloin muodostuu (R)-pantoteenihappoanioni:[36][6]
β-alaniinia syntyy kun L-aspartaatti dekarboksyloituu aspartaatti-1-dekarboksylaasilla (EC 4.1.1.11):[36][6]
- L-aspartaatti → β-alaniini + CO2
Antagonistit[muokkaa | muokkaa wikitekstiä]
Pantoteenihapolla on antagonisteja, jotka estävät sen toimintaa eri tavoin, kuten ottamalla sen paikan entsyymeissä, mutta suorittamatta sen toimintoja.[4] Eräs antagonisti on ω-metyylipantoteenihappo, jossa on pantoteenihapon hydroksimetyyliin (–CH2OH) liittynyt metyyli (–CH(CH3)OH).[37] Aine estää pantototenaattikinaasin (EC 2.7.1.33) toimintaa, jolloin pantoteenihappo ei fosforyloidu 4'-fosfopanteteiiniiksi ja muunnu edelleen koentsyymi A:ksi. Muita antagonisteja ovat destio-CoA, jossa koentsyymi A:n tioli on korvautunut OH-ryhmällä, ja hopantoteenihappo (CAS 18679-90-8), jossa (R)-pantoteenihapon β-alaniini on korvautunut GABA:lla – ketjussa on siis amidisidoksen jälkeen ylimääräinen metyleeni (–CH2–).[4]
-
 Hopantoteenihappo
Hopantoteenihappo -
 ω-Metyylipantoteenihappo
ω-Metyylipantoteenihappo


Nimet ja historia[muokkaa | muokkaa wikitekstiä]
Vuoden 1930 kesäkuussa L. C. Norris ja A. T. Ringrose saivat ruokintakokeissaan aikaan kananpojilla osin pellagran tapaisia oireita, kuten rupista ihottumaa jaloissa ja päässä. He esittivät oireiden johtuvan jonkin tuntemattoman vitamiinin puutteesta – jonkin muun kuin tuolloin vielä huonosti tunnetun "B2-" eli "G-vitamiinin" tai pellagraa ehkäisevän "P-P-tekijän" puutteesta.[38][39] Nämä tunnetaan nykyään vastaavasti nimillä riboflaviini ja niasiini.[40] Vuonna 1939 puuttuva aine varmentui pantoteenihapoksi.[41]
Vuoden 1930 joulukuussa yhdysvaltalainen Roger John Williams (eli vuosina 1893–1988) esitti hiivankasvatuskokeidensa pohjalta olevan olemassa toistaiseksi tuntematon hiivojen kasvuun vaikuttava B-vitamiini.[42][43] Vuonna 1933 jatkotutkimuksissa R. J. Williams ja kollegat eristivät pantoteenihapon epäpuhtaana tutkiessaan hiivojen kasvuun vaikuttavia ravintoaineita. He tunnistivat aineen olevan muun muassa karboksyylihappo. He myös nimesivät aineen: pantoteenihappo tulee kreikan sanasta pantothen (πάντοθεν), joka tarkoittaa "kaikkialta", sillä he arvelivat kokeidensa pohjalta ainetta löytyvän lähes kaikkialta luonnosta. He eristivät ainetta homeesta, naudan maksasta ja kananmunan valkuaisesta.[44][45]
Vuonna 1940 R. J. Williams ja Randolph T. Major (1901–1976) syntetisoivat ensimmäisinä pantoteenihapon.[46][39][47]
Lähteet[muokkaa | muokkaa wikitekstiä]
- GF Combs et al: The vitamins: fundamental aspects in nutrition and health. 3. painos. Elsevier Academic Press, 2008. ISBN 9780121834937.
Viitteet[muokkaa | muokkaa wikitekstiä]
- ↑ a b c Scientific opinion on the safety and efficacy of pantothenic acid (calcium D-pantothenate and D-panthenol) as a feed additive for all animal species based on a dossier submitted by Lohmann animal health: pantothenic acid for all animal species. EFSA Journal, 2011, 9. vsk, nro 11, s. 2409. doi:10.2903/j.efsa.2011.2409. Artikkelin verkkoversio. (Arkistoitu – Internet Archive)
- ↑ a b c d e f M Eggersdorfer et al: ”Vitamins”, Ullmann's Encyclopedia of Industrial Chemistry, s. 123–130. American Cancer Society, 2000. ISBN 9783527306732. Teoksen verkkoversio.
- ↑ a b S Budavari, M O'Neil, A Smith: The Merck index, s. 434. 12. painos. Chapman & Hall Electronic Pub. Division, 2000. ISBN 9781584881292.
- ↑ a b c d e f g h i j k l m n o p q r Combs, s. 346–351
- ↑ a b Pantothenic Acid Drugs.com. Viitattu 13.3.2019.
- ↑ a b c d e f g h i j k l m n o p q r J Zempleni et al: Handbook of vitamins, s. 290–304. 4. painos. Taylor & Francis, 2007. ISBN 9780849340222.
- ↑ a b c d e MA Marahiel, R Finking, HD Mootz: 4′-Phosphopantetheine transfer in primary and secondary metabolism of Bacillus subtilis. Journal of Biological Chemistry, 2001, 276. vsk, nro 40, s. 37289–37298. PubMed:11489886. doi:10.1074/jbc.M103556200. ISSN 0021-9258. Artikkelin verkkoversio.
- ↑ Suomalaiset ravitsemussuositukset 2014. 5. painos. Valtion ravitsemusneuvottelukunta, 2018. ISBN 9789524538015. Teoksen verkkoversio.
- ↑ Scientific opinion on dietary reference values for pantothenic acid. EFSA Journal, 2014, 12. vsk, nro 2, s. 2. doi:10.2903/j.efsa.2014.3581. ISSN 1831-4732. Artikkelin verkkoversio. (Arkistoitu – Internet Archive)
- ↑ Institute of Medicine (US) standing committee on the scientific evaluation of dietary reference intakes, Institute of Medicine (US) subcommittee on interpretation and uses of dietary reference intakes: Using the Adequate Intake for Nutrient Assessment of Groups. National Academies Press, 2000. Teoksen verkkoversio (viitattu 19.3.2019).
- ↑ Dietary reference values for nutrients summary report. EFSA Supporting Publications, 2017, 14. vsk, nro 12, s. 72, 74. doi:10.2903/sp.efsa.2017.e15121. Artikkelin verkkoversio.
- ↑ Overview on tolerable upper intake levels as derived by the Scientific Committee on Food (SCF) and the EFSA panel on Dietetic Products, Nutrition and Allergies (NDA) (pdf) EFSA. 2018. Viitattu 6.3.2019.
- ↑ AL Welsh: Lupus erythematosus: treatment by combined use of massive amounts of panthothenic acid and vitamin E. A.M.A. Archives of Dermatology and Syphilology, 1954, 70. vsk, nro 2, s. 181–198. doi:10.1001/archderm.1954.01540200041004. ISSN 0096-5979. Artikkelin verkkoversio.
- ↑ PC Fry, HM Fox, HG Tao: Metabolic response to a pantothenic acid deficient diet in humans. Journal of Nutritional Science and Vitaminology, 1976, 22. vsk, nro 4, s. 339–346. PubMed:1011047. ISSN 0301-4800. Artikkelin verkkoversio.
- ↑ a b C Gopalan: The 'burning-feet' syndrome. The Indian Medical Gazette, 1946, 81. vsk, nro 1, s. 22–26. PubMed:21025573. ISSN 0019-5863. Artikkelin verkkoversio.
- ↑ a b DJ Lanska: The discovery of niacin, biotin, and pantothenic acid. Annals of Nutrition and Metabolism, 2012, 61. vsk, nro 3, s. 246–253. PubMed:23183297. doi:10.1159/000343115. ISSN 0250-6807. Artikkelin verkkoversio.
- ↑ SW Bibile et al: Pantothenol and the burning feet syndrome. The British Journal of Nutrition, 1957, 11. vsk, nro 4, s. 434–439. PubMed:13489153. doi:10.1079/BJN19570065. ISSN 0007-1145. Artikkelin verkkoversio.
- ↑ a b c MA Kurian, SJ Hayflick: Pantothenate kinase-associated neurodegeneration (PKAN) and PLA2G6-associated neurodegeneration (PLAN): review of two major neurodegeneration with brain iron accumulation (NBIA) phenotypes. International review of neurobiology, 2013, 110. vsk, s. 49–71. PubMed:24209433. doi:10.1016/B978-0-12-410502-7.00003-X. ISSN 0074-7742. Artikkelin verkkoversio.
- ↑ Finto: MeSH: pigmentoitunut pallidusrappeuma finto.fi. Viitattu 8.8.2021.
- ↑ Combs, s. 450–451
- ↑ Combs, s. 449
- ↑ Combs, s. 528–560
- ↑ a b c d e f ME Ensminger et al: The concise encyclopedia of foods & nutrition, s. Ensminger, "Chapter 6 F". CRC Press, 1995. ISBN 9780849344558.
- ↑ Combs, s. 56–57
- ↑ Combs, s. 62
- ↑ S Fanali et al: ”Analysis of vitamins by liquid chromatography”, Liquid chromatography: applications, s. 484. Elsevier Science, 2013. ISBN 9780124158061. doi:10.1016/C2010-0-68910-3.
- ↑ Combs, s. 39
- ↑ a b The Merck index, s. 255. 12. painos. Chapman & Hall Electronic Pub. Division, 2000. ISBN 9781584881292.
- ↑ CG Acevedo-Rocha et al: Microbial cell factories for the sustainable manufacturing of B vitamins. Current Opinion in Biotechnology, 2019, 56. vsk, s. 18–29. doi:10.1016/j.copbio.2018.07.006. Artikkelin verkkoversio.
- ↑ B Shen: Polyketide biosynthesis beyond the type I, II and III polyketide synthase paradigms. Current Opinion in Chemical Biology, 2003, 7. vsk, nro 2, s. 285–295. PubMed:12714063. doi:10.1021/bk-2007-0955.ch011. ISSN 1367-5931. Artikkelin verkkoversio.
- ↑ RD Süssmuth, A Mainz: Nonribosomal peptide synthesis-principles and prospects. Angewandte Chemie International Edition, 2017, 56. vsk, nro 14, s. 3770–3821. doi:10.1002/anie.201609079. ISSN 1433-7851. Artikkelin verkkoversio.
- ↑ KE Bushley, BG Turgeon: Phylogenomics reveals subfamilies of fungal nonribosomal peptide synthetases and their evolutionary relationships. BMC Evolutionary Biology, 2010, 10. vsk, nro 26. PubMed:20100353. doi:10.1186/1471-2148-10-26. ISSN 1471-2148. Artikkelin verkkoversio.
- ↑ JR Lai, A Koglin, CT Walsh: Carrier protein structure and recognition in polyketide and nonribosomal peptide biosynthesis. Biochemistry, 2006, 45. vsk, nro 50, s. 14869–14879. doi:10.1021/bi061979p. ISSN 0006-2960. Artikkelin verkkoversio.
- ↑ DL Nelson, MM Cox: Lehninger principles of biochemistry, s. 530. 5. painos. W.H. Freeman, 2008. ISBN 9780716771081. Teoksen verkkoversio.
- ↑ M Quick, L Shi: The sodium/multivitamin transporter (SMVT): a multipotent system with therapeutic implications. Vitamins and hormones, 2015, 437. vsk, nro 98, s. 63–100. PubMed:25817866. doi:10.1016/bs.vh.2014.12.003. ISSN 0083-6729. Artikkelin verkkoversio.
- ↑ a b c d e f R Leonardi, S Jackowski: Biosynthesis of pantothenic acid and coenzyme A. EcoSal Plus, 2007, 2. vsk, nro 2. PubMed:26443589. doi:10.1128/ecosalplus.3.6.3.4. ISSN 2324-6200. Artikkelin verkkoversio.
- ↑ WB Bean et al: Pantothenic acid deficiency induced in humans subjects. Journal of Clinical Investigation, 1955, 34. vsk, nro 7 Pt 1, s. 1073–1084. PubMed:14392222. doi:10.1172/JCI103156. ISSN 0021-9738. Artikkelin verkkoversio.
- ↑ AT Ringrose, LC Norris: The occurrence of a pellagrous-like syndrome in chicks. Science, 1930, 71. vsk, nro 1851, s. 643–643. PubMed:17839940. doi:10.1126/science.71.1851.643. ISSN 0036-8075. Artikkelin verkkoversio.
- ↑ a b GD Novelli: Metabolic functions of pantothenic acid. Physiological Reviews, 1953, 33. vsk, nro 4, s. 525–543. PubMed:13100068. doi:10.1152/physrev.1953.33.4.525. ISSN 0031-9333. Artikkelin verkkoversio.
- ↑ Combs, s. 23–29
- ↑ LC Norris, GF Heuser, MB Gillis: The need for pantothenic acid and an unidentified factor in reproduction in the domestic fowl. The Journal of Nutrition, 1942, 23. vsk, nro 2, s. 153–163. doi:10.1093/jn/23.2.153. ISSN 0022-3166. Artikkelin verkkoversio.
- ↑ RJ Williams, EM Bradway: The further fractionation of yeast nutrilites and their relationship to vitamin b and wildiers' “bios”. Journal of the American Chemical Society, 1931, 53. vsk, nro 2, s. 783–789. doi:10.1021/ja01353a051. ISSN 0002-7863. Artikkelin verkkoversio.
- ↑ FF Nord, CH Werkman: ”The chemistry and biochemistry of pantothenic acid”, Advances in Enzymology and Related Areas of Molecular Biology, s. 255. Wiley, 1943. ISBN 9780470122488. Teoksen verkkoversio.
- ↑ RJ Williams et al: “Pantothenic acid,” a growth determinant of universal biological occurrence. Journal of the American Chemical Society, 1933, 55. vsk, nro 7, s. 2912–2927. doi:10.1021/ja01334a049. ISSN 0002-7863. Artikkelin verkkoversio.
- ↑ DR Davis, ML Hackert, LJ Reed: Roger J. Williams 1893-1988 (pdf) 2008. National academy of sciences. Arkistoitu 12.9.2015.
- ↑ Randolph T. Major, Roger J. Williams: The structure of pantothenic acid. Science, 1940, 91. vsk, nro 2358, s. 246–246. PubMed:17831185. doi:10.1126/science.91.2358.246. ISSN 0036-8075. Artikkelin verkkoversio.
- ↑ Vitamin B complex American Chemical Society. Arkistoitu 7.7.2017. Viitattu 17.3.2019.
Aiheesta muualla[muokkaa | muokkaa wikitekstiä]
- Linus Pauling Institute: pantothenic acid (englanniksi)
- Human Metabolome Database: Pantothenic acid
- KEGG: Pantothenic acid (englanniksi)
- Liber Herbarum II: Luettelo pantoteenihappoa sisältävistä kasveista
| ||||||||||||||||||||||||||||||