Arotundra



Arotundra oli jääkauden aikainen kitukasvuinen kasvillisuusvyöhyke, jossa kasvoi lähinnä ruohoja, heiniä ja saroja sekä joskus pensaita ja puita. Kitukasvuisuus johtui kuivuudesta, kylmyydestä, ilman hiilidioksidin alhaisuudesta ja eläinten laidunnuksesta. Aron/aavikon ja tundran kasveja kasvoi mosaiikkina. Lunta oli talvella yleensä vähän. Arotundran kasvit kasvoivat tuulen kuljettamassa lössissä ikiroudan päällä. Maa oli kalkkipitoista. Kun ilma oli kuivaa, soita oli harvemmassa kuin nyt ja maa paikoin suolaista. Arotundraa on usein sanottu mammuttiaroksi, koska siellä viihtyivät mammutit, hevoset, peurat ja muut jääkautiset eläimet.[1]
Arotundran tyyppikasveja olivat muun muassa maruna, savikka ja lapinvuokko. Kosteammilla arotundran alueilla kasvoi myös puita tavallisesti paikallisina metsinä. Varsinkin jääkauden lauhoina, kosteina interstadiaaleina arotundran paikalla oli laajalti avointa puistomaista metsäaroa. Arotundra katosi jääkauden päättyessä, kun ikirouta suli ja virtaavien jäätiköiden kulutuksen aiheuttama lössin muodostus loppui. Pohjoisessa arotundra korvautui jääkauden lopussa vaivaiskoivikolla. Nykyisten Keski-Aasian arojen muun muassa Mongoliassa on katsottu muistuttavan jääkautista arotundraa. Suomessa rantojen, tunturien ja harjujen kasvillisuuden, muun muassa ruijanesikko-ryhmän, katsotaan yleensä olevan arotundran jäänteitä.
Kasvit[muokkaa | muokkaa wikitekstiä]

Arotundralla kasvoi aro- ja tundrakasveja rinta rinnan.[2][3][4][5][6][7][8][9][10]
Tämä johtui kuivuudesta ja kylmyydestä. Arotundra oli tundraa kuivempi ja melkein aina nykyisiä aroja kylmempi avoin matala kasvillisuustyyppi, jossa kasvoi monin paikoin niittymäisesti lähinnä ruohoja, heiniä, saroja ja kuivilla alueilla marunaa,[11][12][13] kuivilla paikoilla savikkaa[11][14] sekä joskus pensaita[15] ja paikoin puita.
Arotundrakasvillisuus oli yleensä hyvin harvaa[12] ja muistutti nykyistä kuivaa aroa, puoliaavikkoa tai kuivaa tundraa. Monin paikoin tundrakasvit olivat enemmän vuoriston kuin tundran kasveja. Arotundran kuivimpien alueiden kasvit olivat niin sanottuja pioneerilajeja.
Esimerkiksi Baltiasta tavatun arotundran kasvit olivat tundrakasveja, kuten vaivaiskoivu (Betula nana) ja lapinvuokko, kuivuudesta kieliviä, kuten maruna ja savikka ja suolasta kieliviä, kuten salsola-pensas[9]
Tyypillinen kylmän arotundran kasvi on karuilla mailla lapinvuokko (Dryas octopetala).[16] Kuivuutta, kylmyyttä taas luonnehti kuuramaruna (Artemisia frigida). Erilaiset tuppaina kasvavat nurmikat ja heinät muun muassa itunata (Festuca vivipara) luonnehtivat arotundraa. tunturisarake (Kobresia myosuroides) oli myös yleinen varsinkin Beringiassa. Kosteammilla alueilla kasvoi erilaisia pajulajeja. Tundramaisessa Beringiassa ja Itä-Siperiassa Selaginella.[12] Arotundran kasvit voidaan laajasti katsoen jakaa ensinnäkin metsän ja niityn mesofyytteihin, joita oli lauhkeita termofiilisiä sekä mikrotermisiä eli tundrakasveja.[17] Ensimmäisin kuuluu muun muassa hanhentatar (Persicaria maculosa), toiseen tunturilieko (Diphasiastrum alpinum) ja mähkä (Selaginella selaginoides). Toiseksi oli mesofyyttisiä arokasveja lähinnä jauhosavikka (Chenopodium album) ja varsinkin sinisavikka (C. glaucum) ja punasavikka (C. rubrum). Kserofyyttisiä arokasveja olivat Ephedra distachya ja marunat (Artemisia).[17] Nämä kuivakkokasvit veivät ilmaston kylmettyä tilaa vaivaiskoivulta.[18] Kuivinta kautta ja aluetta luonnehtivat suolakkokasvit, muun muassa Bassia prostrata[17][19]
Harvoista puuvartisista kasveista kuivuutta luonnehti lähinnä kataja[20] ja etelämpänä mänty, kylmyyttä vaivaiskoivu[21] ja vaivaispaju.
Eläimistö[muokkaa | muokkaa wikitekstiä]
Eläimet olivat yleensä aron ja tundran eläimiä, mutta myös metsäeläimiä esiintyi.[22] Arotundralla eli eläinlajeja kaksi kertaa niin paljon kuin nykyisellä tundralla.[23] Esimerkiksi mammuttia[24] suojasi paksu turkki.[25] Kylmään sopeutumiseen liittyi myös mammutin pienet korvat ja lyhyt kärsä. Tyypillisiä olivat biisonit,[26] myskihärät,[26] mammutit, peurat[27] ja hevoset,[2][15][26] ja myös villasarvikuono[26][28] Kasvinsyöjäeläimet hyötyivät arotundran lössistä, koska saivat siitä tarvitsemiaan kivennäisiä. Napakettu viihtyi kylmemmillä alueilla. Sen sijaan saigaa tavattiin harvoin Pohjois-Siperiassa. Luolahyeena, luolaleijona, luolakarhu, susi ja ihminen saalistivat näitä eläimiä.[28] Pienimpiä arotundran eläimiä olivat sopulit ja lapinmyyrä.
Ilmasto, maaperä[muokkaa | muokkaa wikitekstiä]

Arotundralla vallitsi mannerilmasto, jossa vuotuiset lämpötilan vaihtelut olivat suuria. Oli usein hyvin kylmää ja/tai tuulista.[29] Kesä oli lyhyt ja talvi pitkä. Tuuli kuljetti jäätikön ääreistä pölyä kauas jäätikön reunasta. Arotundran maaperä oli laajalti kalkkipitoisen pölyn, lössin lannoittamaa ja painanteissa monesti suolaantunut.[30][31] Niinpä arotundran monesti kalkkipitoinen[32] maaperä oli korkeintaan lievästi hapan, neutraali tai emäksinen/alkalinen. Zimovin mukaan arotundra ei eronnut välttämättä tavallisesta tundrasta niinkään kesä- ja sademäärän perusteella, vaan runsaan lössilannoituksen takia kasvavat arokasvit haihduttivat tundrakasveja enemmän, ja kuivattivat maata.[33] Typestä oli laajalti pulaa.[34][35] Niinpä lössi ja eläinten laidunnuksen vaikutus selittää arotundran runsaan kasvillisuustuoton tundraan verrattuna.[36]
Kun maaperästä haihtui kosteutta, maannos oli kuiva ja hapekas, ei nykyisen tyyppinen soinen turvemaannos tai muuten kostea. Tällöin kivennäisiä irtosi nopeasti rapautuvassa maannoksessa, mikä tarjosi kasveille paljon ravinteita.[35] Tämä mahdollisti kosteata tundraa suuremman biomassan tuotannon, mikä elätti suuria ruohonsyöjäeläimiä, jotka tuottivat kasveille ravinteita kuollessaan, ulostaessaan ja virtsatessaan.[35] Muun muassa mammutit tarvitsivat kivennäisiä, esimerkiksi kalsiumia ja natriumia.[37][38] Eläimet myös mylläsivät maata kulkiessaan. Niinpä arotundran kasvilajiston uskotaan muokkautuneen laidunnuksen mukaan, niin kuin Jakutian nykyisillä arolaikuillakin.[39]
Guthrien mukaan Beringian arotundra muistutti jääkaudella sataneen lössin takia laajalti enemmän kosteaa ei-hapanta tundraa, kuin nykyistä hapanta kosteaa tundraa.[40] Ei-happamassa kosteassa tundrassa eli MNT:ssä (MNT = moist non-acidic tundra) maannos oli kalsiumrikas, hienojakoinen ja melko hapan. Runsasravinteinen MNT kasvaa enemmän myrkyttömiä, kalsium- ja muuten ravintopitoisia kasveja.[40]
Arotundraa luonnehti myös ikirouta,[41] jonka pintakerros suli kesällä kyllin, jotta kasvit saattoivat kasvaa[42][43] mutta eläimet kulkea ikiroudan päällä. Monet kryogeeniset prosessit olivat kuivemmilla mantereilla alueilla harvinaisia, yleensä pakkasen aiheuttamia halkeamia.[44] Arotundra ulottui nykyisen lauhkean vyöhykkeen leveysasteilla Euroopasta Keski-Aasiaan, Siperiaan ja Alaskaan asti[2] ja tästä itäänkin Arktisten saarten jäättömille alueille.[45] Varsinkin jääkauden lämpimämpien vaiheiden aikana monet suuret kasvinsyöjäeläimet viihtyivät arotundralla.[46]
Alue[muokkaa | muokkaa wikitekstiä]
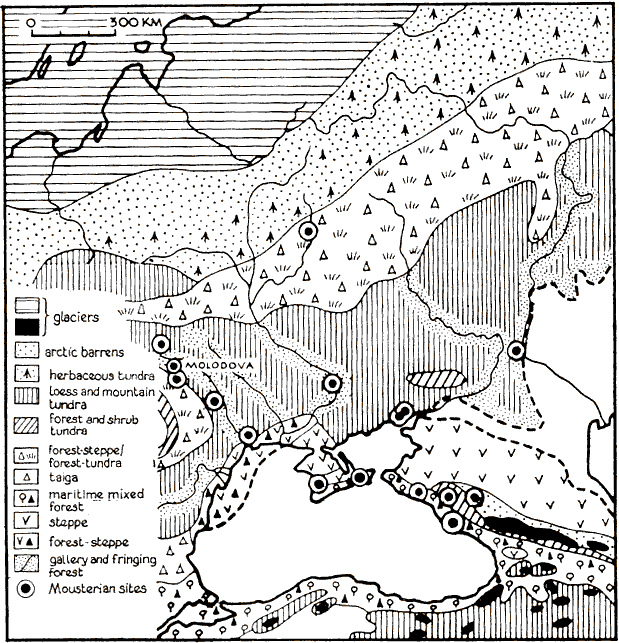
Laajasti sanottuna arotundra kattoi jääkauden huippukaudella Etelä-Ranskasta Itä-Siperiaan ulottuvan noin 40–50 leveysasteen pohjoispuolisen alueen[42][47][48][49] jossa esiintyi ikiroutaa[50][51][52] ainakin epäjatkuvana[53] ja oli ainakin paikoin lössiä[54][55] ja mammutteja.[56] Puustoa oli niukasti,[42] lähinnä jokilaaksoissa ja vuorilla. Tällä alueella oli hyvin erilaisia ympäristöjä polaariaavikosta, tundrasta, arosta[42] ja metsästä aavikkoon. Oli myös näiden välimuotoja pensastundraa, metsätundraa ja metsäaroa sekä metsäaron metsätundran välimuotoa.
Nimitys arotundra liitetään varsinkin Ukrainan ja Keski-Siperian alueelle, lännessä oli kosteampaa. Läntisen Venäjän arotundra jaetaan yleensä neljään vyöhykkeeseen: pohjoinen tundramainen alue, Valko-Venäjän periglasiaalinen metsäaro, Ukrainan periglasiaalinen aro.[57] Tundramaisella alueella oli paljasta maata ja yrttikasveja kasvavaa tundraa,[58] aroa, metsiä joissa lehtikuusta, mäntyä, koivua ja lisäksi hajanaisia suolakkokasviyhdyskuntia.[57] Oli sekä aro- että tundrakasvistoja,[11] yrttejä, marunoita, savikkakasveja,[59] koivissa paikoissa runsaasti yrttikasveja.[12] Eteläisempi periglasiaalinen metsäaro oli yhä arktinen[60] päätellen sopulin esiintymisestä, ja siellä kasvoi aromaista kasvistoa, lehtikuusta, mäntyä ja koivua sekä tundraa joissain kylmissä painumissa.[57] Niinpä tämä alue oli metsätundraa ja metsäaroa.[58] Aromaiseen kasvillisuuteen kuului marunoita, yrttejä, savikkaa ja efedraa.[12] Lössiä ja lössihiuetta 400–600 metriä merenpinnasta asti.[12] Tätäkin vyöhykettä voidaan pitää lämpötilaltaan ainakin pensastundrana[61] tai metsätundrana[62] ja Ukrainassa sen pohjoisosissa saattoi kasvaa korkeaa ruohoa ja puita.[63]
Tämän arktisen vyöhykkeen eteläpuolella oli periglasiaalinen aro, jossa saattoi olla suuria lehtimetsäsaarekkeita jokien varsilla ja vuoristoissa muun muassa Etelä-Uralilla.[57] Arokasvillisuus oli marunaa, savikkakasveja, yrttikasveja ja suolakkokasveja.[12] Muuten periglasiaalisen aron vuoristoissa oli tundraa, alavilla alueilla pelkkää metsää ja pensastundraa.[58] Tämä vyöhyke ulottui idässä Etelä-Jakutiaan asti.[57][64] Se oli Länsi-Aasiassa harvakasvista, mutta Baikaljärven itäpuolella puita oli enemmän ja kasvillisuus peittävää.[12] Eläimistö oli pohjoisella periglasiaalisella arolla osin boreaalista, muun muassa hevosia, muuten sielläkin oli tyypillinen mammuttieläimistö.[65] Jääkauden lopun ajan eli nuoremman dryaskauden arotundralle oli ominaista lyhyt kuuma kesä, pitkä kylmä talvi.[11] Ilmasto muistutti etäisesti nykyajan Jakutian ilmastoa.
Etäisesti arotundraa muistuttavia alueita on Keski-Jakutian aroilla, niin kuin myös Janan, Indigirkan ja Kolyman jokialtaissa, joissa kasvaa metsätundraa.[11] Kasvukausi oli lyhyt, kosteus ei riittänyt metsälle tundralla ja kosteus arolla. Jääkauden arotundralla aron ja tundran kasviyhdyskunnat kasvoivat erillään. Nykyajan Venäjän aroilla on havaittavissa arktinen kasvuaika talvella, tundra-aika varhain keväällä, metsäaika myöhäiskeväällä ja aroaika kesällä.[11] Myös tundralta löytyvät vastaavat vuodenajat. Niinpä aro- ja tundrakasvillisuutta saattoi kasvaa suunnilleen samoilla seuduilla.[11]
Erilaisia arotundria[muokkaa | muokkaa wikitekstiä]
Monesti Pohjois-Siperian ja Alaskan jääkautinen arotundra oli käytännössä kasviyhdyskuntien mosaiikki.[66] Niissä, esimerkiksi ylhäällä vuoristossa kasvoi kuivatundrakasvillisuutta, hieman alempana niityillä tunturisaraketta ja/tai kunnailla lapinvuokkoa,[67] etelärinteiden keskiosissa tuppaina marunan ja tupasheinien[68] tyyppisiä arokasveja, jokilaaksoissa kuivia ja kosteita niittyjä ja jokivarsilla märän ympäristön kasveja, joissain paikoissa dyynejä, suolakkokasveja jne.[69][70][71]
Jokilaaksot olivat jääkaudella Pohjois-Siperiassa luultavasti kuivempia ja suottomampia kuin nyt. Ylempänä tundra oli enemmän nykyisen ylämaan tundran tyyppistä.[72]
Euraasiassa Kazakstanissa Semipalatinskin lähellä oli lämpimänä jääkauden kautena alemmilla, loivilla jokilaaksoterasseilla aroa, ylemmällä havupuita, jokilaakson terassien välisissä painumissa suota ja koivua/haapaa, samoin kuin jokilaaksojen tulvatasangoilla, Kylmänä kautena alemmilla jokiterasseilla kasvoi koivu-kuusimetsän ja arokasvillisuuden mosaiikkia, ylimmällä terassilla vaivaiskoivua ja koivua sekä aroa, terassien välissä oli järviä ja soita, tulvatasangoilla kasvoi tyrnipensaita.[73][74]
Alaskasta Sewardin niemimaalta löydetty Kitluk-joelta jääkautinen tuliperäisen tuhkan alle hautautunut paleomaannos, jossa näyttää olleen tasapainossa oleva kasviyhdyskunta, missä kasvoi sammalen päällä tunturisaraketta ja tundralle tyypillisiä ruohoja, ja jossa monista arotundran alueista poiketen oli maassa paljon typpeä ja fosforia.[75][76]
Beringian arotundran tyypillisiä ovat nadat Festuca, nurmikat Poa botryoides, Poa stepposa, Poa arctostepporum, pahtanurmikka (Poa Glauca), Calamagrostis purpurascens, Helicotrichon krylovii, Koeleria cristata, Koeleria asiatica ja Elytrigia-sukuisia vehnäruohoja. Sarojen joukossa oli monenlaisiin ympäristöihin sopeutuneita, esimerkiksi aito arokasvi Carex diriuscula, niittyarojen kasveja Carex obtusata, kalliosara (Carex rupestris), tuppaina kasvavia saroja muun muassa rinnejalkasara (Carex pediformis), Carex filifolia, Carex rossii, Carex aenea, Carex patasata jne. Kuivaan ja kuivaan/kylmään sopeutuneita lajeja olivat marunat (Artemisia), hanhikit (Potentilla), kurjenherneet (Astragalus), keulankärjet (Oxytropis).[77] Tunturikurjenherneen (Astragalus alpinus) ja idänkaulankärjen (Oxytropis campestris) tyyppiset herneen sukuiset kasvit olivat tyypillisiä jäätikön läheiselle alueelle, jossa sademäärä oli pieni ja maaperä neutraali,[78] ja pulaa typestä. Näissä hernekasveissa on paljon eläimille sopivaa valkuaista.
Ajallinen ja paikallinen vaihtelu[muokkaa | muokkaa wikitekstiä]
Mosaiikkimaisen[79][80] arotundran kasvillisuus ja kosteus vaihtelivat muun muassa eri alueilla, leveysasteilla ja aikoina. Parhaimmillaan arotundra oli subarktista metsäaroa tai puistomaista aluetta, jolla puita kasvoi ruohoalueen seassa hieman samaan tapaan kuin Afrikan savannilla tai Kanadan Aspen parklandissa. Nykyään uskotaan puita kasvaneen joissain painumissa esimerkiksi Alppien pohjoispuolella[81] ja jopa Norjan jäättömissä jokilaaksoissa.[82] Arotundraa on sanottu usein myös mammuttiaroksi.[2][83] Täsmällisesti sanoen mammuttiaro kattoi myös ne alueet, joilla ei esiintynyt tundrakasvillisuutta kuten Etelä-Ukrainassa. Hyvin kuivalla aavikkoa muistuttavalla arotundralla kasvoi savikan sukuisia kasveja. Ephedra distachyan[84] tyyppinen yrtti luonnehti kuivaa arotundran niittyä. Kuivempi arotundra suosi enemmän hevosta kuin mammuttia.
Jääkauden maksimin aikaan arotundralla oli laajojen dyynikenttien keskellä kosteita painumia ja jokilaaksoja, joissa oli muun niittyä ja kosteikkokasvillisuuttakin. Yleensä painumat olivat suolaisia, ja niissä viihtyivät runsastuottoiset suolaa sietävät kasvit. Painumiin saattoi virrata vuorilta tai pulputa lähteestä makeaa vettä. Lössialueella kasvoi lyhytkasvuista aroheinää.[85]
Vuorten rinteillä saattoi kasvaa pensaikoita tai metsiä, samoin jokilaaksoissa. Melko lämpimissä, kosteissa suolapainumissa saattoi olla kasvillisuudeltaan runsastuottoisia alueita. Vuorten juurilla olevat alueet minne valui vuorilta jäätiköiden ja lumien sulamisvesiä, saattoivat olla tuottoisia. Arotundran tuottoisuudesta ja kyvystä elättää mammutin tapaisia suuria eläimiä on kiistelty tieteessä pitkään, mutta keskustelu siitä on nyttemmin jäämässä taka-alalle. Esimerkiksi mammutin tiedetään löydöistä päätellen viihtyneen jokivarsien lähellä, ei kuivimmilla alueilla.
Viime jääkauden keskivaiheilla oli suuria jäätiköitä, jotka kuivattivat ilmaa, mutta maan akseli oli kallellaan ja tuotti nykyistä lämpimämpiä kesiä Pohjois-Siperiaan. Pohjois-Siperiassa arotundra oli aromaisimmillaan noin 48 000–35 000 radiohiilivuotta sitten,[86][87] eli silloin oli lämmintä ja kuivaa.[88] Etelän arokasvit muun muassa harjasvehnä (Agropyron cristatum) vaelsivat pohjoiseen.[89]
Lämpimin huippu oli itäisimmässä Siperiassa noin 38 000 radiohiilivuotta sitten,[90] jolloin siellä lehtikuusimetsät valtasivat alaa tundralta.[91] Monet Pohjois-Siperian mammuttilöydöt ajoittunevat tähän lämpökauteen.[92] Mutta jääkauden kylmimmän jakson aikaan Pohjois-Siperian kasvillisuus muistutti lähinnä kuivaa tundraa, koska aidot arokasvit olivat kadonneet.[93] Mutta kuiva tundra vuorotteli kosteiden niittyjen kanssa.[94]
Melko lämmintä oli uudestaan jääkauden lopussa noin 18 000–12 500 radiohiilivuotta sitten.[86][88]
Puolassa näkyy jääkauden lopun ajan arotundran vaihdos paju-savikkatyyppisestä koivu-artemisiatyyppiseen[95] luultavasti ilmaston lämpenemisen ja/tai kostumisen takia.
Vertailu nykyisiin kasvillisuustyyppeihin[muokkaa | muokkaa wikitekstiä]
Siperia[muokkaa | muokkaa wikitekstiä]
Jäänteitä arotundrasta saattaa olla joillain arktisilla alueilla muun muassa Wrangelin saarella. Luoteis-Siperiassa on nykyäänkin paikoin aroja ja tundra-aroja.[96][97] Arotundraa on usein vertailtu Jakutian Sartan-joen aroalueisiin, vaikka nykyään tutkijat eivät usko sen vastaavan jääkautista ympäristöä. Samankaltainen alue on muun muassa Tšuraptšan aro Keski-Jakutiassa.[98] Varsinkin Jakutian jokivarsien kuivilla, jyrkillä ylärinteillä oleva lyhytruohoinen arokasvillisuus tuo mieleen mammuttiaron.[99] Keski-Jakutian Lena-joen laakson ja ikiroudan sulamispainumien kasvillisuus muistuttaa luultavasti mammuttien elinympäristöä aroineen ja soisine suolaniittyineen.[100][101][102][103] Luoteis-Siperian pohjoisboreaalisen ja tundran arot jaetaan ruohomätäsaroihin, kuivahkoihin niittyihin ja harvaan kasvillisuuteen.[104]
Altai, Keski-Aasia[muokkaa | muokkaa wikitekstiä]
Arotundra muistutti kuivimmillaan laajalti lähinnä Keski-Aasian kuivia aroja,[105] puoliaavikoita ja aavikoitakin,[106]. Maruna ja savikka ovat näiden alueiden siitepölyissä yleisiä, ja runsas maruna liittyy niillä aroilla runsaisiin heinäkasveihin.[107] Näille ovat monesti tyypillisiä yleisympäristöön nähden rikas eläimistö ja painumien ja vuorenrinteiden runsas kasvillisuus ja hevoseläinten asuminen. Maruna on näillä aroilla runsainta, kun sademäärä on 200–700 mm.[108]
Näitä on Mongoliassa, Ubsunurin alueella, Džungariassa, Gobissa ja Tiibetin Arjin Shanin luonnonsuojelualueella. Varsinkin Altai-Sajanin seudun alueen siitepölyt muistuttavat jääkautisia. [109] Altai-Sajanin Khindiktig-Khol’-järven allas (Mongun-Taigan massiivi, Tuvanin tasavalta) ja Bertekin painanne Ukokin tasangolla Altain tasavallassa saattoivat muistuttaa jääkauden ympäristöjä.[110] Ukok-Saliugemin laakson, Khar us Nuurin ja Ubsunurin altaan peura ja saiga muistuttavat ehkä jääkautisia.[111] Kohoton-Khorgonin järvillä sataa 294 mm vuodessa, tammikuussa on −26 °C ja heinäkuussa vain 9.4 °C.[112]
Myös Tiibetin ylänköä voi pitää arotundraa muistuttavana, koska siellä elää niukalla kasvillisuudella suuria eläimiä muun muassa jakkeja,[113] kesä on viileä ja talvi kylmä.
Amerikassa, Grönlannissa[muokkaa | muokkaa wikitekstiä]
Arotundraa löytyy myös Kalliovuorilta, muun muassa vuorten välisistä laaksoista. Myös Nebraska Sandhillsin dyynialueen siitepölyt ovat lähellä jääkautisia .Maruna liittää arotundran Pohjois-Amerikan lyhytheinäpreeriaan, joka on biisonien luonnollinen elinympäristö. Berezovkan mammutin kasvijäänteet tuovat mielen Mongolian arojen joenvarsiniityt ja silloiset järvien lähistöt.[114]
Alaskan Kobuk-joen varren dyynit muistuttavat jääkautisia dyynialueita, ja niiden lähellä on monenlaisia kasviyhdyskuntia.
Alaskan pingojen etelärinteitä on myös verrattu arotundrakasvillisuuteen. Varsinkin Brooks-vuorten pohjoispuolisia kalsiumrikkaita tundria on pidetty ainakin jossain määrin jääkautista arotundraa muistuttavina.[115] Joissain järvien ja jokien ympäröimissä laaksoissa saattoi olla arktisia keitaita, jollainen Ellesmerensaaren Lake Hazen nykyään on. Jääkautisen tyyppisen laidunnuksen aiheuttamia muutoksia kuvastaa esimerkiksi Grönlannin ala-arktisen/subarktisen sisämaan pajua kasvavan aroalueen muutos niittynurmikkaruohikoksi.[116] Länsi-Grönlannin ala-arktinen tai subarktinen Kangrelussuaqin vaivaiskoivu-vaivaispaju-tunturisarake-heinäkasvimosaiikki tuo mieleen arotundran, jossa on myös suolajärviä.
Muualla[muokkaa | muokkaa wikitekstiä]
Götlannin kalkkipitoisilla alvareilla kasvaa muun muassa päivännoutoa ja nataa, niin kuin arotundralla oletetaan kasvaneen.[117][118]
Hieman arotundran tyyppisiä alueita on Suomessa nykyään muun muassa Käsivarren Lapin suurtuntureilla, Kuusamossa, rannikoilla, tienpientareilla, kalliokedoilla ja harjuilla[119] sekä lehdoissakin.
Nykyisissä kasvillisuustyypeissä näkyy varsinkin pohjoiselle arotundralle tyypillistä mosaiikkimaisuutta. Esimerkiksi Lapissa on lämpimämpää etelärinteillä, mutta kylmempää pohjoisrinteillä. Mäen päällä saattaa olla viisi astetta lämpimämpää kuin laaksossa, mutta hieman ylempänä olevassa rinteen painanteessa viisi astetta kylmempää kuin laaksossa.[120] Toisaalta melko syvälumiset lumenviipymät suosivat ruohojen kasvua, matalalumiset varpujen kasvua.
Kasvimaantieteellisistä rajoista[muokkaa | muokkaa wikitekstiä]

Arotundra oli monien arvioiden mukaan toisaalta kylmää, toisaalta kuivaa. Niinpä monet nykyiset kasvillisuustyypit ovat joko liian lämpimiä tai kosteita arotundraksi.[121] Venäjän arotundralla saattoi kasvaa puita kylmistä talvista huolimatta, koska ilmasto oli kuiva. Koivu (Betula pubescens) saattaa mannermaisessa ilmastossa kestää tammikuussa -40 °C keskilämpötilan ja heinäkuussa 12 °C. Meri-ilmastossa sama koivu kestää vain -12 °C keskilämpötilan tammikuussa. Samoin mänty (Pinus sylvestris) kestää meri-ilmastossa -12 °C keskilämpötilan tammikuussa ja 12 °C heinäkuussa, mutta vastaavasti -40 °C/14 °C kuivassa mannerilmastossa.[122] Vuotuisen kuukauden keskilämpötilan vaihtelu eli mantereisuus oli ikiroudan rajan pohjoispuolella laajalti 40–50 °C,[123] mikä merkitsi laajoilla alueilla heinäkuun lämpötiloja 10–19 °C ja tammikuun -21 – -60 °C.[124] Valko-Venäjän metsäaro-metsätundravyöhykkeellä lämpötila oli tammikuussa noin -22 – +30 °C, kesällä -14 °C.[124] Siellä satoi 200–350 mm[123] ja kuukausien suurin lämpötilaerotus oli -35 – -45 °C.[123] Tämä vastasi korkeintaan Siperian pohjoista taigaa kesälämpötilojen puolesta, ja talvilämpötilojen puolesta jopa Etelä-Siperian keskimmäistä tai eteläistä lehtikuusi-mäntytaigaa.[125] Tällöin suuressa osassa arotundraa pystyi lämpötilan puolesta kasvamaan puita, ja kuivuus oli suurempi ongelma puiden kasvulle.
Monien kasvillisuusarvioiden mukaan Pohjois-Aasian arotundran lössialueilla olisi laajoilla alueilla satanut 40–60 % siitä mitä nyt eli 300 mm mutta maaperätutkimusten mukaan alle 200 mm.[126] Kasvitutkimusten mukaan sademäärä oli usein välillä 250–350 mm.[123] Tundran tapauksessa tämä olisi saattanut tarkoittaa sitä, että arotundra muistutti enemmän Itä-Siperian tundraa, joissa sataa 250–350 mm jolloin sadanta/haihduntasuhde on 1,1–1,2 eli sama kuin metsissä.[125] Ilmasto on luultavasti ollut paljon tätä kuivempi. Nykyisillä eteläisillä ”Artemisia-aavikoilla” sataa vain 150 mm ja puoliaavikoilla 350 mm.[125] Näissä haihdunnan ja sadannan suhde on luokkaa 0,15–0,5.[125] Tämä ennustaa kuivimmalle arotundralle sademäärää 40–120 mm.
Arotundran katoaminen[muokkaa | muokkaa wikitekstiä]
Arotundran häviäminen liittyi tavalla tai toisella jääkauden aikaisten eläinten tuhoon. Arotundra hävisi jääkauden päättyessä ilmaston kostuessa ja lämmetessä. Lämpeneminen sulatti jäätiköt, mikä lopetti lössin synnyn. Myös ikirouta suli laajalti. Lämpeneminen ja kostuminen metsitti ja soisti maastoa. Kosteus huuhtoi maasta kivennäisravinteita pois. Niinpä maaperä happani ja kasvilajivalikoima köyhtyi ja muuttui. Ensin ilmestyi pensaita, sitten aluksi varvikkoisia vaivaiskoivumetsiä syrjäyttämään heinäkasvit. Kanervat ja varvut yleistyivät heinäkasvien sijaan. Kun kosteus huuhtoi maaperän mineraalit pois, eläinten oli vaikeampi saada kivennäisiä.[127] Jääkautiset myskihärän tyyppiset eläimet olivat yleensä tottuneet melko kuivaan ympäristöön. Kosteammat talvet saattoivat olla vastasyntyneelle vasikoille vaikeampia, koska näillä ei ollut kostealta ja kylmältä suojaavaa turkkia. Lämpö ja kosteus edistivät loisten elämää. Talvinen lumen syveneminenkin saattoi aiheuttaa ongelmia, vaikka esimerkiksi myskihärän, hevosen ja biisonin tiedetäänkin löytävän ravintoa melko syvästä lumesta.[33]
Arotundran kasveja[muokkaa | muokkaa wikitekstiä]
Joitain arotundran kasveja:
-
 Marunat (Artemisia)
Marunat (Artemisia) -
 Sarat (Cyperaceae)
Sarat (Cyperaceae) -
 Heinät (Poaceae)
Heinät (Poaceae) -
 Hanhikit (Potentilla)
Hanhikit (Potentilla) -

-
 Vaivaiskoivu (Betula nana)
Vaivaiskoivu (Betula nana) -
 Pajut (Salix)
Pajut (Salix) -
 Lakka (Rubus chamaemorus)
Lakka (Rubus chamaemorus)








Katso myös[muokkaa | muokkaa wikitekstiä]
Lähteet[muokkaa | muokkaa wikitekstiä]
- Barnosky, C. W. & Velichko, A. A. & Wright, H. E.: Late-quaternary environments of the Soviet Union. London: Longman, 1984. ISBN 0-582-30125-4.
- Björklund, Tom & Vuokko, Seppo: Mammutin aika, elämää jääkaudella. Helsinki: Tammi, 2009. ISBN 978-951-31-4113-4.
- David M. Hopkins & John V. Matthews & Charles E. Schweger: Paleoecology of Beringia. New York: Academic Press, 1982. ISBN 978-0-12-355860-2.
- Eronen, Matti: Jääkausien jäljillä. Ursan julkaisuja 43. Helsinki: Ursa, 1991. ISBN 951-9269-59-2.
- Fagan, Brian: Pitkä kesä. Suomentanut Saarinen, Osmo. Helsinki: Ajatus, 2008. ISBN 978-951-20-7595-9.
- Frederick Hadleigh West: American Beginnings, the Prehistory and Palaecology of Beringia. University of Chicago Press, 1996. ISBN 0-226-89399-5.
- Frenzel, Burkhard & Pecsi, Marton & Velichko, Andrej Alekseevich (päätoim.): Atlas of Paleoclimates and Paleoenvironmants of the Northern Hemisphere - Late Pleistocene-Holocene. Stuttgart, Jena, New York., Bremerhaven: Geographical Researh Institute, Hungarian Academy of Sciences, Gustav Fischer Verlag, PANGAEA, 1992. ISBN 3-437-30685-X. Teoksen verkkoversio (viitattu 16.7.2019). doi:10013/epic.37670.d001
- Frenzel, Burkhard & Pecsi, Marton & Velichko, Andrej Alekseevich: Atlas of Paleoclimates and Paleoenvironments of the Northern Hemisphere (digitized version). PANGAEA, 2001. Teoksen verkkoversio (viitattu 16.7.2019). Pictures - Paleoatals
- Hallanaro, Eeva-Liisa & Pylvänäinen, Marja & From, Stella: Pohjois-Euroopan luonto : löytöretki monimuotoisuuteen. Helsinki: Edita, 2002. ISBN 92-893-0636-X.
- Hoffecker, John F. & Elias, Scott A.: Human ecology of Beringia. New York: Columbia University Press, 2007. ISBN 978-0-231-13060-8.
- Koivisto, Marjatta: Jääkaudet. Helsinki: WSOY, 2004. ISBN 951-0-29101-3.
- Kurtén, Björn: Jääkausi. Helsinki: WSOY, 1972. ISBN 951-0-00149-X.
- Kurtén, Björn: Kuinka mammutti pakastetaan. Suomentanut Irma Rostedt. Helsinki: Tammi, 1982. ISBN 951-30-5627-9.
- Lappalainen, Iiris: Suomen luonnon monimuotoisuus. Helsinki: Suomen ympäristökeskus, Edita, 1998. ISBN 951-37-2380-1.
- Lister, Adrian & Bahn, Paul: Mammutit: Jääkauden jättiläiset. (Mammoths – giants of the ice age, 2007.) Esipuhe: Jean M. Untinen-Auel. Suomentanut Arja Hokkanen. Helsinki: WSOY, 2000. ISBN 951-0-25064-3.
- Taipale, Kalle : Tulivuorista jääkausiin : Suomen maankamaran kehitys , s. 302–303. Helsinki : WSOY , 1991 . ISBN 951-0-16048-2.
- Soffer, Olga: The Upper Paleolithic of the Central Russian Plain. Orlando, Fla: Academic Press, 1985. ISBN 0-12-654271-6.
- Walter, K. M. & Edwards, M. E. & Gross G. et al.: Thermokarst Lakes as a Source of Atmospheric CH4 During the Last Deglaciation. Science, 2007, 318. vsk, nro 5850, s. 633–636. doi:10.1126/science.1142924. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- Wiik, Kalevi: Eurooppalaisten juuret. Jyväskylä: Atena Kustannus Oy, 2002. ISBN 951-796-250-9.
Lähteet[muokkaa | muokkaa wikitekstiä]
- ↑ Kurtén, Björn: Jääkauden Euroopassa komeita eläimiä Tiede -lehti. 1.1.2004. Viitattu 14.7.2019.
- ↑ a b c d Pirrus, R. & Raukas, A.: Late-glacial stratigraphy in Estonia. Proceedings of the Estonian Academy of Sciences, Geology, 1996, 45. vsk, nro 1, s. 34–45. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Sauer, Jonathan D.: Plant Migration: The Dynamics of Geographic Patterning in Seed Plant Species, s. 148. University of California Press, 1988. Teoksen verkkoversio (viitattu 14.7.2019).
- ↑ Muller, S. & Tarasov, P. E. & Andreev, A. A. & Diekmann, B.: Late Glacial to Holocene environments in the present-day coldest region of the Northern Hemisphere inferred from a pollen record of Lake Billyakh, Verkhoyansk Mts, NE Siberia. Climate of the Past, 2009, s. 73–84. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Tarasov, P. E. & Bezrukova, E. V. & Krivonogov, S. K.: Late Glacial and Holocene changes in vegetation cover and climate in southern Siberia derived from a 15 kyr long pollen record from Lake Kotokel. Climate of the Past, 2009, nro 5, s. 285–295. doi:10.5194/cp-5-285-2009. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Vasari, Yrjö & Kuznetsov, Oleg L. & Lavrova, Nadezhda B. & Shelekhova, Tatyana S. & Vasari, Annikki: Alinlampi, a Late-Glacial site in the northern Karelian Republic. Annales Botanici Fennici, 2007, 44. vsk, s. 42–55. Finnish Zoological and Botanical Publishing Board. ISSN 0003-3847. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Yelovicheva, Yadviga & Sanko, Aleksandr: Palynostratigraphy of the Poozerie Glaciation (Vistulian) in Belarus. Kwartalnik Geologiczny, 1999, 43. vsk, nro 2, s. 203–212. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Kihnoa, Kersti & Saarsec, Leili & Amon, Leeli: Late Glacial vegetation, sedimentation and ice recession chronology in the surroundings of Lake Prossa, central Estonia. Estonian Journal of Earth Sciences, 2011, 60. vsk, nro 3, s. 147–158. doi:10.3176/earth.2011.3.03. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ a b Older Dryas chronozon. Proceedings of the Estonian Academy of Sciences, Geology 1996, 1996, 45. vsk, nro 1, s. 40. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Henriksen, M., Mangerud, J., Matiouchkov, A., Murray, A. S., Paus, A. & Svendsen, J. I.: Intriguing climatic shifts in a 90 kyr old lake record from northern Russia. Boreas, , 37. vsk, nro 1, s. 20–37. doi:10.1111/j.1502-3885.2007.00007.x. ISSN 0300-9483. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ a b c d e f g Velichko 1984, s. 187.
- ↑ a b c d e f g h Frenzel 1992, s. 55.
- ↑ Zazulaa, Grant D. & Froese, Duane G. & Elias, Scott A. & Kuzmina, Svetlana & Mathewes, Rolf W.: Arctic ground squirrels of the mammoth-steppe: paleoecology of Late Pleistocene middens (∼24 000–29 450 14C yr BP), Yukon Territory, Canada. Quaternary Science Reviews, 2007, 26. vsk, nro 7–8, s. 979–1003. doi:10.1016/j.quascirev.2006.12.006. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Koivisto 2004, s. 186.
- ↑ a b Lister & Bahn s. 74.
- ↑ Tulivuorista jääkausiin, Taipale, Kalle, s. 302–303
- ↑ a b c Velichko 1984, s. 173. Alkup. artikkeli Late pleistocene vegetation history, V.P. Grichuk
- ↑ Gey, Valery & Saarnisto, Matti & Lunkka, Juha Pekka & Demidovc, Igor: Mikulino and Valdai palaeoenvironments in the Vologda area, NW Russia. Global and Planetary Change, 2001, 31. vsk, nro 1–4, s. 347–366. doi:10.1016/S0921-8181(01)00129-1. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Vinogradova, Olena & Kiosak, Dmytro: The Holocene Palaeovegetation of South-Western Ukraine: A Review. Bretschneider-online.it, 2007. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Tolksdorf, Johann Friedrich & Turner, Falko & Kaiser, Knut & Eckmeier, Eileen & Stahlschmidt, Mareike & Housley, Rupert A. & Breest, Klaus & Veil, Stephan: Multiproxy Analyses of Stratigraphy and Palaeoenvironment of the Late Palaeolithic Grabow Floodplain Site, Northern Germany. Geoarchaeology, 2013, 28. vsk, nro 1, s. 50–65. doi:10.1002/gea.21429. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Bos, Johanna A.A. & Dambeck, Rainer & Kalis, Arie J. & Schweizer, A. & Thiemeyer, Heinrich: Palaeoenvironmental changes and vegetation history of the Upper Rhine Graben (southwestern Germany) since the Lateglacial. Geologie en Mijnbouw, 2008, 87. vsk, nro 1, s. 67–90. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Soffer 1985, s. 153, 155, 190.
- ↑ Fagan 2008, s. 63.
- ↑ Kurten 1972, s. 72, 75.
- ↑ Kurten 1972, s. 75.
- ↑ a b c d Kurten 1972, s. 77.
- ↑ Kurten 1972, s. 78.
- ↑ a b Eronen 1991, s. 202.
- ↑ Pitkä kesä, Brian Fagan 2008, s. 41.
- ↑ Björklund & Vuokko 2009, s. 94.
- ↑ Kasvit muuttuvassa metsäluonnossa, Antti Reinikainen & Raisa Mäkipää..., ISBN 951-31-1963-7, Tammi Helsinki 2000, s. 26.
- ↑ Koivisto 2004, s. 187.
- ↑ a b Zimov, S. A. & Chuprynin, V. I. & Oreshko, A. P. & Chapin III, F. S. & Reynolds, J. F. & Chapin, M. C.: Steppe-Tundra Transition: A Herbivore-Driven Biome Shift at the End of the Pleistocene. The American Naturalist, 1995, 146. vsk, nro 5, s. 765–789. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Reinikainen 2000, s. 26.
- ↑ a b c Hoffecker & Elias 2007, s. 58, s. 59, Alkuperäinen lähde Zimov et al 1995
- ↑ Zimov, S.A. & Zimov, N.S. & Tikhonov, A.N. & Chapin III, F.S.: Mammoth steppe: a high-productivity phenomenon. Quaternary Science Reviews, 2012, 57. vsk, nro 4, s. 26–45. doi:10.1016/j.quascirev.2012.10.005. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Leshchinskiy, S.V.: The Late Pleistocene beast solonetz of Western Siberia: “mineral oases” in mammoth migration paths, foci of the Palaeolithic man’s activity. La Terra Degli Elefanti–The World of Elephants: Proceedings of the First International Congress, 2001. Department of Palaeontology & Historical Geology, Tomsk State University, Tomsk, Russia. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Haynes, Gary: Mammoth landscapes: good country for hunter-gatherers. Quaternary International, 2006, 142–143. vsk, s. 20–29. doi:10.1016/j.quaint.2005.03.002. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Werger, Marinus J.A. & van Staalduinen, Marja A. (Eds.): ”Antopogenic impacts”, Plant and Vegetation 6 – Eurasian Steppes. Ecological Problems and Livelihoods in a Changing World, s. 388, 389. Springer, 2012. ISBN 978-94-007-3885-0. Teoksen verkkoversio (viitattu 14.7.2019). (englanniksi)
- ↑ a b Hoffecker & Elias 2007, s. 57
- ↑ Frenzel 1992, s. 49, 53.
- ↑ a b c d Hallanaro 2002, s. 35.
- ↑ Swanson, David: Biogeographical Evidence for the Grass (Poaceae) Species of Pleistocene Beringian Lowlands. Arctic, 2006, 59. vsk, nro 2, s. 191–200. National Park Service. doi:10.14430/arctic341. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Frenzel 1992, s. 120.
- ↑ Eronen 1991, s. 199.
- ↑ Björklund & Vuokko 2009, s. 51.
- ↑ Lister & Bahn, s. 27.
- ↑ Adams, Jonathan: Global land environments since the last interglacial Oak Ridge National Laboratory, TN, USA. 1997. Viitattu 14.7.2019.
- ↑ Adams, J.M. & Faure, H.: Review And Atlas Of Palaeovegetation: Preliminary land ecosystem maps of the world since the Last Glacial Maximum Oak Ridge National Laboratory, Oak Ridge, TN 37831, USA. 10.2.1997. Viitattu 14.7.2019.
- ↑ Eronen 1991, s. 193, 199.
- ↑ Koivisto 2004, s. 205.
- ↑ Frenzel 1992, s. 120, s. 51, 49, 53, 57, 59, 61, 63.
- ↑ Velichko 1984, s. 76.
- ↑ Velichko 1984, s. 94.
- ↑ Walter, Edwards ja Gross, s. 633–636.
- ↑ Frenzel 1992, s. 60-61.
- ↑ a b c d e Velichko 1984, s. 176, 177.
- ↑ a b c Dons Maps, vegetation Zones of Ukraine (alkup Soffer, World at 18000 BP) Don's Maps. Viitattu 14.7.2019.
- ↑ Velichko 1984, s. 187
- ↑ Velichko 1984, s. 18, s. 176, s. 177
- ↑ Wiik 2002, s. 68.
- ↑ Kurten 1972, s. 66–67
- ↑ Fagan 2008, s. 48
- ↑ Frenzel 1992, s. 59, 57.
- ↑ Frenzel 1992, s. 61.
- ↑ Zazula, Grant D. & Froese, Duane G. & Westgate, John A. & Farged, Catherine La & Mathewes, Rolf W.: Paleoecology of Beringian bpackratQ middens from central Yukon Territory, Canada. Quaternary Research, 2005, 63. vsk, s. 189–198. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Kienast, Frank & Schirrmeister, Lutz & Siegert, Christine & Tarasov, Pavel: Palaeobotanical evidence for warm summers in the East Siberian Arctic during the last cold stage. Quaternary Research, 2005, 63. vsk, nro 3, s. 283–300. doi:10.1016/j.yqres.2005.01.003. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Zazula, Grant D. & Froese, Duane G. & Schweger, Charles E. & Mathewes, Rolf W. & Beaudoin, Alwynne B. & Telka, Alice M. & Harington, C. Richard & Westgate, John A.: Ice-age steppe vegetation in east Beringia. Nature, 2003, 423. vsk, nro 6940, s. 603. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Zazula, Grant D. & Schweger, Charles E. & Beaudoin, Alwynne B. & McCourtc, George H.: Macrofossil and pollen evidence for full-glacial steppe within an ecological mosaic along the Bluefish River, eastern Beringia. Quaternary International, 2006, 142–143. vsk, s. 2–19. doi:10.1016/j.quaint.2005.03.010. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Yurtsev, Boris A.: The Pleistocene “Tundra-Steppe” and the productivity paradox: the landscape approach. Quaternary Science Reviews, 2001, 20. vsk, nro 1–3, s. 165–174. doi:10.1016/S0277-3791(00)00125-6. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Kirillova, Irina V. & Argant, J. & Lapteva, E.G. & Korona, O.M. & Plicht, J. van der & Zinovyev, E.V. & Kotov, А.А. & Chernova, O.F. & Fadeeva, Е.О. & Baturina, O.A. & Kabilov, M.R. & Shidlovskiy, F.K. & Zaninag, O.G.: The diet and environment of mammoths in North-East Russia reconstructed from the contents of their feces. Quaternary International, 2016, 406. vsk, nro Part B, s. 147–161. doi:10.1016/j.quaint.2015.11.002. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Kienast, Frank & Schirrmeister, Lutz & Siegert, Christine & Tarasov, Pavel: Palaeobotanical evidence for warm summers in the East Siberian Arctic during the last cold stage. Quaternary Research, 1.5.2005, 63. vsk, nro 3, s. 283–300. doi:10.1016/j.yqres.2005.01.003. ISSN 0033-5894. Artikkelin verkkoversio.
- ↑ Rutter, Nat W. & Velichko, A. A. & Dolukhanov, P.M. & Rutter, N. W. & Catto, N.R.: Quaternary of northern Eurasia: Late Pleistocene and Holocene landscapes, stratigraphy and environments. Quaternary international, 1997, 41/42. vsk, s. 128–129. ISSN 1040-6182. (englanniksi)
- ↑ Kremenetski, C.V. & Tarasov, P.E. & Cherkinsky, A.E.: The latest pleistocene in southwestern Siberia and Kazakhstan. Quaternary International, 1997, 41–42. vsk, s. 125–134, 127. doi:10.1016/S1040-6182(96)00044-4. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Goetcheus, Victoria G. & Birks, Hilary H.: Full-glacial upland tundra vegetation preserved under tephra in the Beringia National Park, Seward Peninsula, Alaska. Quaternary Science Reviews, 2001, 20. vsk, nro 1–3, s. 135–147. doi:10.1016/S0277-3791(00)00127-X. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Höfle, Claudia & Edwards, Mary E. & Hopkins, David M. & Mann, Daniel H. & Chien-Lu Ping: The Full-Glacial Environment of the Northern Seward Peninsula, Alaska, Reconstructed from the 21,500-Year-Old Kitluk Paleosol. Quaternary Research, 2000, 53. vsk, nro 2, s. 143–153. doi:10.1006/qres.1999.2097. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Hoffecker & Elias 2007, s. 30.
- ↑ Paus, Aage: Late Weichselian vegetation, climate and floral migration at Eigebakken, South Rogaland, Southwestern Norway. Review of Palaeobotany and Palynology, 1989, 61. vsk, nro 3–4, s. 177–203. doi:10.1016/0034-6667(89)90032-8. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Zimov 1981.
- ↑ Hofreiter, Michael & Stewart, John: Ecological Change, Range Fluctuations and Population Dynamics during the Pleistocene. Current Biology, 2009, 19. vsk, nro 14, s. R584–R594. doi:10.1016/j.cub.2009.06.030. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Birks, H. John B. & Willis, Katherine J.: Alpines, trees, and refugia in Europe. Plant Ecology & Diversity, 2008, 1. vsk, nro 2, s. 147–160. doi:10.1080/17550870802349146. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Kaaro, Jani: Tutkimus: Kuuset sinnittelivät Skandinaviassa jääkauden läpi Helsingin Sanomat. 3.3.2012. Viitattu 14.7.2019.
- ↑ Eronen 1991, s. 201, 199.
- ↑ Marjatta Koivisto: Jääkaudet, 2004. s. 187.
- ↑ Kurten 1972, s. 68.
- ↑ a b Kienast, Frank & Schirrmeister, Lutz & Siegert, Christine & Tarasov, Pavel: Palaeobotanical evidence for warm summers in the East Siberian Arctic during the last cold stage. Quaternary Research, 2005, 63. vsk, nro 3, s. 283–300. doi:10.1016/j.yqres.2005.01.003. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Müller, S. & Andreev, A. & Diekmann, B. & Zech, W: Late Pleistocene and Early Holocene Vegetation and Climate History in the Verkhoyansk Mountains (Yakutia, Russia) Derived from Pollen Records. 22. Internationale Polartagung, Jena, Deutschland., 18.–24.09.2005. Alfred Wegener Institut. doi:10013/epic.23910. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ a b Schirrmeister, Lutz & Siegert, Christine & Kuznetsova, Tatyana & Kuzmina, Svetlana & Andreev, Andrei & Kienast, Frank & Meyer, Hanno & Bobrov, Anatoly: Paleoenvironmental and paleoclimatic records from permafrost deposits in the Arctic region of Northern Siberia. Quaternary International, 2002, 89. vsk, nro 1, s. 97–118, s. 116. doi:10.1016/S1040-6182(01)00083-0. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Werger, Marinus J.A. & van Staalduinen, Marja A. (Eds.): ”Antopogenic impacts”, Plant and Vegetation 6 – Eurasian Steppes. Ecological Problems and Livelihoods in a Changing World, s. 378. Springer, 2012. ISBN 978-94-007-3885-0. Teoksen verkkoversio (viitattu 14.7.2019). (englanniksi)
- ↑ West 1996, s. 132.
- ↑ West 1996, s. 133.
- ↑ West 1996, s. 131, 132
- ↑ Sher, Andrei: The Last Glacial Environment in Beringida - an Attempt of Multidisciplinary and Long-distance Comparison: Andrei Sher, Session No. 217 Geology and Paleoecology of the Beringian Subcontinent: To Honor the Career of David M. Hopkins, 2003 Seattle Annual Meeting (November 2–5, 2003) Geological Society of America, 2003 Seattle Annual Meeting. 2–5.11.2003. Viitattu 14.7.2019. (englanniksi)
- ↑ Geel, Basvan & Aptroot, André & Baittinger, Claudia & Birks, Hilary H. & Bull, Ian D. & Cross, Hugh B. & Evershed, Richard P. & Gravendeel, Barbara & Kompanje, Erwin J.O. & Kuperus, Peter & Mol, Dick & Nierop, Klaas G.J. & Pals, Jan Peter & Tikhonov, Alexei N. & Reenen, Guidovan & Tienderen, Peter H. van: The ecological implications of a Yakutian mammoth's last meal. Quaternary Research, 2008, 69. vsk, nro 3, s. 361–376. doi:10.1016/j.yqres.2008.02.004. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Wacnik, Agnieszka & Ralska-Jasiewiczowa, Magdalena & Madeyska, Ewa & Szafer, W.: Late Glacial and Holocene history of vegetation in Gostynin area, central Poland. Acta Palaeobotanica, 2011, 51. vsk, nro 2, s. 249–278. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Hopkins 1982, s. 159.
- ↑ Yurtsev, Boris A.: 9 – Relics Of The Xerophyte Vegetation Of Beringia In Northeastern Asia. Paleoecology of Beringia, 1982, s. 157–177. doi:10.1016/B978-0-12-355860-2.50018-1. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Velichko 1984, s. 115.
- ↑ Latchininsky, Alexandre V.: Grasshopper Problems in Yacutia (Eastern Siberia, Russia) Grasslands – Natural Conditions, Climate and Vegetation. Journal of Orthoptera Research, 1995, nro 4, s. 29–34. doi:10.2307/3503455. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Troeva, E.I. & Isaev, A.P. & Cherosov, M.M. & Karpov, N.S. (Eds.): ”Luku 3 Vegetation of Yakutia: Elements of Ecology and Plant Sociology & luku 5 Insect Impact on Vegetation”, The Far North: Plant Biodiversity and Ecology of Yakutia, s. s. 155, 313. Springer, 2010. ISBN 978-90-481-3773-2. Teoksen verkkoversio (viitattu 14.7.2019).
- ↑ Troeva, E. I. & Cherosov, M. M.: Transformation of Steppe Communities of Yakutia Due to Climatic Change and Anthropogenic Impact, s. 371–396. Springer, 2012. ISBN 978-94-007-3885-0. Teoksen verkkoversio (viitattu 14.7.2019).
- ↑ Shahid, Shabbir A. & Abdelfattah, Mahmoud A. & Taha, Faisal K. (Eds.): ”The New Map of Soil Salinity and Regularities in Distribution of Salt-Affected Soils in Russia”, Developments in Soil Salinity Assessment and Reclamation : Innovative Thinking and Use of Marginal Soil and Water Resources in Irrigated Agriculture, s. 107. Springer, 2013. ISBN 978-94-007-5683-0. Teoksen verkkoversio (viitattu 14.7.2019).
- ↑ Werger, Marinus J.A. & Staalduinen, Marja A. van: ”Steppe types of Yakutia”, Eurasian Steppes. Ecological Problems and Livelihoods in a Changing World, s. 373– s. 378, s. 374. Springer, 2012. ISBN 978-94-007-3885-0. Teoksen verkkoversio (viitattu 14.7.2019).
- ↑ Hopkins 1982, s. 158.
- ↑ Björklund & Vuokko 2009, s. 93.
- ↑ Velichko 1984, s. 117.
- ↑ Hongyan Liu & Yi Wang & Yuhong Tian & Jiangling Zhu & Hongya Wang: Climatic and anthropogenic control of surface pollen assemblages in East Asian steppes. Review of Palaeobotany and Palynology, 24.1.2006, 138. vsk, nro 3, s. 281–282. Department of Ecology, College of Environmental Sciences, Peking University. Artikkelin verkkoversio (pdf). Viitattu 14.7.2019. (englanniksi)
- ↑ Liu Hongyan & Liu Kan & Wei Fangling: Artemisia pollen-indicated steppe distribution in southern China during the Last Glacial Maximum. Journal of Palaeogeography, 2013, 2. vsk, nro 2, s. 297–305. Department of Ecology and Ministry of Education Key Laboratory of Earth Surface Processes, College of Urban and Environmental Sciences. doi:10.3724/SP.J.1261.2013.00032. Artikkelin verkkoversio (pdf). Viitattu 14.7.2019. (englanniksi)
- ↑ Blyakharchuka, T.A. & Wright, H.E. & Borodavko, P.S. & Knaap, W.O. van der & Ammann, B.: Late Glacial and Holocene vegetational changes on the Ulagan high-mountain plateau, Altai Mountains, southern Siberia. Palaeogeography, Palaeoclimatology, Palaeoecology, 2004, 209. vsk, nro 1–4, s. 259–279. doi:10.1016/j.palaeo.2004.02.011. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Current palynological pattern of steppe-tundra at the Altai highlanddepressions Ivan Strelkov, Ekaterina Nosevich, Mikhail Amosov, and Kirill Chistyakov https://presentations.copernicus.org/EGU2020/EGU2020-7245_presentation.pdf
- ↑ Řičánková, Věra Pavelková & Robovský, Jan & Riegert, Jan: Ecological Structure of Recent and Last Glacial Mammalian Faunas in Northern Eurasia: The Case of Altai-Sayan Refugium. PLoS ONE, 13.1.2014, 9. vsk, nro 1. PubMed:24454791. doi:10.1371/journal.pone.0085056. ISSN 1932-6203. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ https://web.archive.org/web/20181210154624/https://www.globalspecies.org/weather_stations/climate/537/278
- ↑ Suttie, J.M. & Reynolds, S.G.: ”Chapter VIII – China case study 3: Pastoral systems, change and the future of the grazing lands in Tibet – Tashi Nyima”, Transhumant Grazing Systems in Temperate Asia. Rome: Food and Agriculture Organization of the United Nations, 2003. ISBN 92-5-104977-7. Teoksen verkkoversio (viitattu 14.7.2019). (englanniksi)
- ↑ Suttie, J.M. & Reynolds, S.G.: ”Chapter III – Mongolia case study 1: Studies on long-distance transhumant grazing systems in Uvs and Khuvsgul aimags of Mongolia, 1999-2000 – B. Erdenebaatar”, Transhumant Grazing Systems in Temperate Asia. Rome: Food and Agriculture Organization of the United Nations, 2003. ISBN 92-5-104977-7. Teoksen verkkoversio (viitattu 14.7.2019). (englanniksi)
- ↑ Walker, D.A & Bockheim, J.G & Chapin III, F.S & Eugster, W & Nelson, F.E & Ping, C.L: Calcium-rich tundra, wildlife, and the “Mammoth Steppe”. Quaternary Science Reviews, 2001, 20. vsk, nro 1–3, s. 149–163. doi:10.1016/S0277-3791(00)00126-8. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Kalaallit Nunaat low arctic tundra World Wildlife Fund WWF. Viitattu 14.7.2019. (englanniksi)
- ↑ Löbel, Swantje & Dengler, Jürgen: Dry grassland communities on southern Öland: phytosociology, ecology, and diversity. Acta Phytogeographica Suecica, 2008, 88. vsk, s. 13–32. Artikkelin verkkoversio. Viitattu 14.7.2019. (englanniksi)
- ↑ Carlsson, Bengt & Maarel, Eddy van der: Structure and dynamics of alvar vegetation on Oland and some related dry grasslands. Uppsala: , 2007. ISBN 978-91-7210-088-6. Teoksen verkkoversio (viitattu 14.7.2019).
- ↑ Lappaleinen 1998, s. 56, 40.
- ↑ Luonto talvella, Sonia Eriksson Hans Heorg Wallentius, Gummerus 1986, ISBN 951-20-2817-4, s. 10
- ↑ Velichko 1984, s. 290.
- ↑ Velichko 1984, s. 278.
- ↑ a b c d Velichko 1984, s. 282.
- ↑ a b Velichko 1984, s. 280, 281.
- ↑ a b c d Velichko 1984, s. 288–289.
- ↑ Velichko 1984, s. 284.
- ↑ Reumer, Jelle W.f. & Vos, John De: Abstracts (3) 2nd International Mammoth Conference. 1999 may 16 – may 20. Arkistoitu 6.7.2009. Viitattu 14.7.2019. (englanniksi)
{kind=link}
Aiheesta muualla[muokkaa | muokkaa wikitekstiä]
- Paleo Data Search National Centers for Environmental Information (NCEI). Viitattu 15.7.2019. (englanniksi)
- Food for Thought for Scientists Hans Krause’s Research Reports . Viitattu 15.7.2019. (englanniksi)